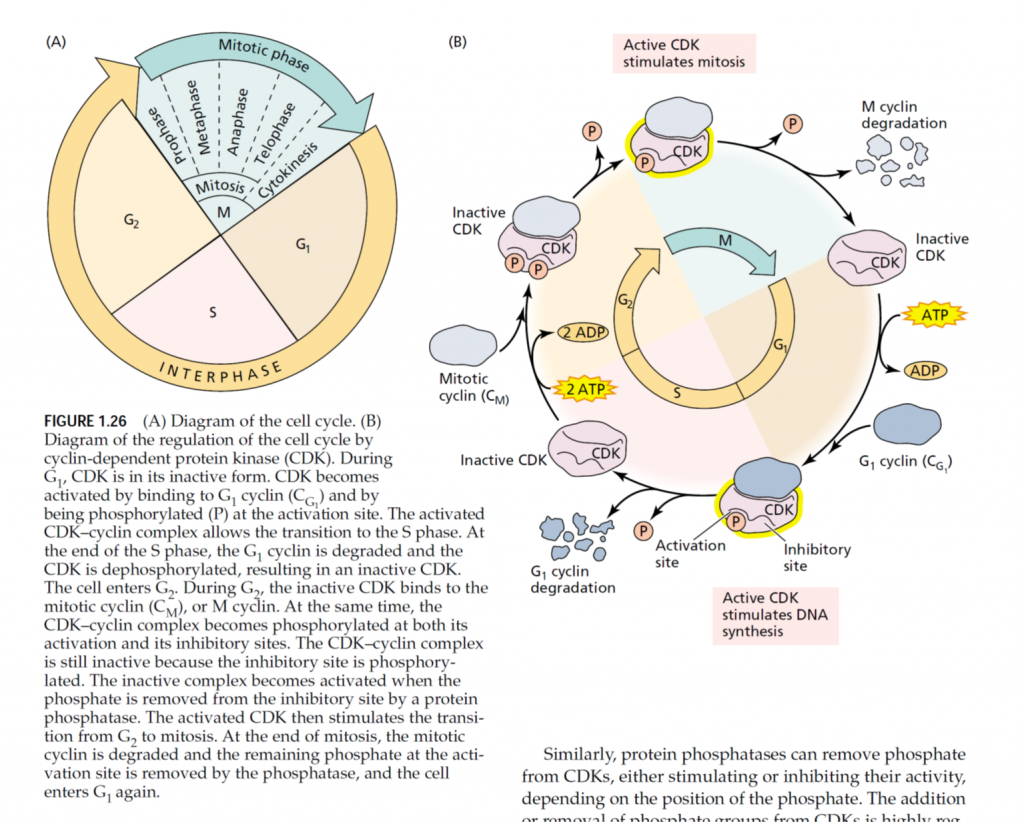
What really sets a Master Gardener apart from other home gardeners is their special training in horticulture. In exchange for their training, individuals who become Master Gardeners volunteer their time, working through their cooperative Extension office, to provide horticultural-related information to their communities.
Master Gardeners are members of the local community who take an active interest in their lawns, trees, shrubs, flowers and gardens. They are enthusiastic, willing to learn and to help others, and able to communicate with diverse groups of people.
bove is an example of a nutrient labels guaranteed analysis. It tells us how much of each element is in the bag at percentage weight by volume (%w/v). This provides us with enough information to establish a reasonably accurate ppm.
Note that analyzing the ppm from fertilizer labels won’t provide 100% accurate ppms. Fertilizers sold worldwide are often only required to be listed accurately to within 0.4%.
Regulations around the world require that NPK.. values be presented somewhat ambiguously. Therefore, listings for the same nutrient may appear to vary on a country-by-country basis. For example, when looking at our labels guaranteed analysis you will find note that it states;
Available Phosphate (P2O5)……….15.0% Available Potash (K2O)…………..14.0%
This information becomes important when interpreting the guaranteed analysis.
That is, it is important to note that the P and K numbers found on the guaranteed analysis do not always reflect the actual amounts of elemental phosphorous and potassium by %.
With our label, this is the case and P is listed as P2O5 (phosphorous pentoxide) and K is listed as K2O (potassium oxide) percentage.
When phosphorus is listed as P2O5 it is only 43% elemental P and when potassium is listed as K2O it is only 83% elemental K.
Therefore, when this system is in use, a 5-15-14 NPK ratio truly reflects elemental NPK 5-6.45-11.62.
N = 5 P = 15 * 0.43 = 6.45 K = 14 * 0.83 = 11.62
Additionally, other nutrients such as calcium (Ca), magnesium (Mg) and sulfur (S) can be listed in their oxide form (CaO, MgO, SO3) or in elemental form, or both. To convert other nutrient listings that may appear on some labels use these equations.
CaO to Ca multiply by 0.714 MgO to Mg multiply by 0.6031 SO3 to S multiply by 0.4
Percentage Weight by Volume (%w/v)
A simple way of understanding how to convert a %w/v listing found on the guaranteed analysis into grams per litre is by understanding that 1ml of RO water weighs 1gram.
Percentage weight by volume %w/v refers to the total weight of elements contained within a finished concentrate of a given total volume.
For example, 5% of nitrogen added to 1 litre(1000ml) of RO water would mean that there is 50grams of N in the water.
1000 (ml) * 0.05 (5% nitrogen) = 50 (grams of N)
Converting %w/v to ppm and ppm to %w/v
To establish ppm from %w/v you simply need to multiply by 10000. 5% (nitrogen) * 10000 = 50000 (ppm) To establish %w/v from ppm you simply need to divide by 10000. 50000 (ppm) / 10000 = 5% (nitrogen)
To establish the concentration of individual elements in the water, the guaranteed analysis (%w/v) should first be converted into ppm, then multiplied by the usage rate (per litre), then divided by 1000 (ml).
For example, if a nutrient lists 5% nitrogen, when it is used at 5grams per 4 litres it will yield 62.5 ppm of nitrogen per litre.
c.1942 potassium carbonate (K2CO3); c.1950 any one or more of potassium chloride (KCl), potassium sulfate (K2SO4) or potassium nitrate (KNO3).[9][10] Does not contain potassium oxide (K2O), which plants do not take up.[11] However, the amount of potassium is often reported as K2O equivalent (that is, how much it would be if in K2O form), to allow apples-to-apples comparison between different fertilizers using different types of potash.
Potassium oxide (K2O) is an ionic compound of potassium and oxygen. The chemical formula K2O (or simply ‘K’) is used in several industrial contexts: the N-P-K numbers for fertilizers,
The phosphate or orthophosphate ion [PO 4]3− is derived from phosphoric acid by the removal of three protons H+
Lime
Agricultural lime, also called aglime, agricultural limestone, garden lime or liming, is a soil additive made from pulverized limestone or chalk. The primary active component is calcium carbonate..Calcium oxide (CaO), is commonly known as quicklim.e
Calcium carbonate reacts with water that is saturated with carbon dioxide to form the soluble calcium bicarbonate.
CaCO3(s) + CO2(g) + H2O(l) → Ca(HCO3)2(aq)
Agriculture and aquaculture
Agricultural lime, powdered chalk or limestone, is used as a cheap method for neutralising acidic soil, making it suitable for planting, also used in aquaculture industry for pH regulation of pond soil before initiating culture.[54]
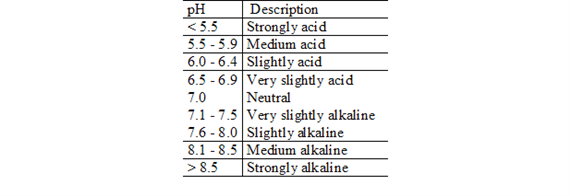
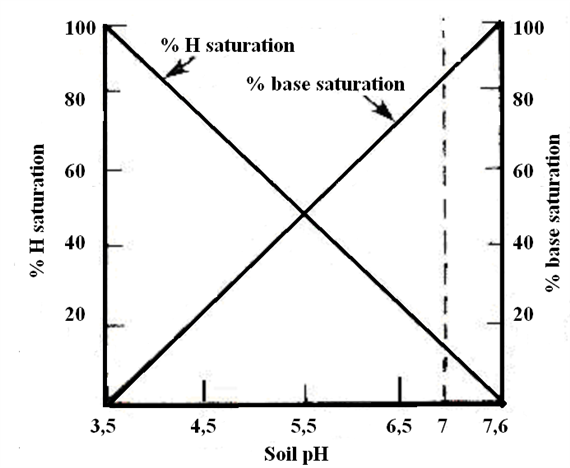
Soil pH defines the relative acidity or alkalinity of the soil solution (Table 1.). The pH scale in natural systems ranges from 0 to 14. A pH value of 7.0 is neutral. Values below 7.0 are acid and those above 7.0 are alkaline, or basic. Many agricultural soils have a soil pH between 5.5 and 6.5.
Soil pH is a measurement of hydrogen ion (H+) activity, or effective concentration, in a soil and water solution. Soil pH is expressed in logarithmic terms, which means that each unit change in soil pH amounts to a tenfold change in acidity or alkalinity. For example, a soil with a pH of 6.0 has 10 times as much active H+ as one with a pH of 7.0.
Descriptive terms commonly associated with certain ranges in soil pH are:
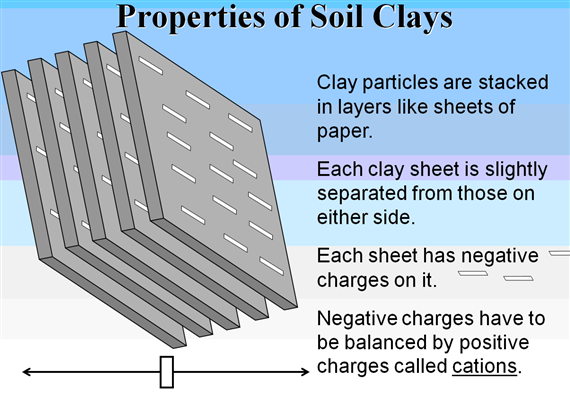
Cation and anion exchange capacity (CEC)
Cation-exchange capacity is defined as the degree to which a soil can adsorb and exchange cations.on surface with negative charge.
Sources of negative charge:
The main source of charge on clay minerals is isomorphous substitution which confers permanent charge on the surface of most layer silicates.
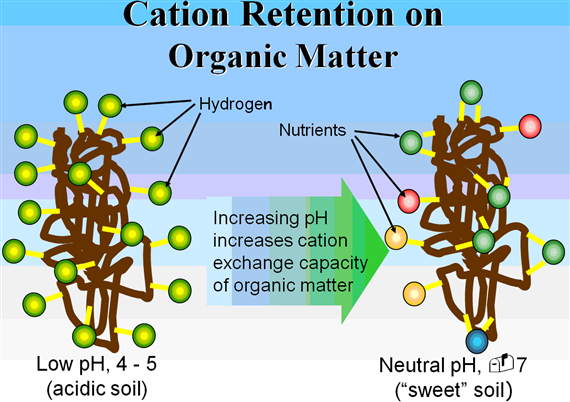
Ionization of hydroxyl groups on the surface of other soil colloids and organic matter can result in what is describes as pH dependent charges-mainly due to the dependent on the pH of the soil environment. Unlike permanent charges developed by isomorphous substitution, pH-dependent charges are variable and increase with increasing pH.
Presence of surface and broken – edge -OH groups gives the kaolinite clay particles their electronegativity and their capacity to absorb cations. In most soils there is a combination of constant and variable charge. Cation-a positively charged ion There are two types of cations, acidic or acid-forming cations, and basic, or alkaline-forming cations. The Hydrogen cation H+ and the Aluminum cation Al+++ are acid-forming.
The positively charged nutrients that we are mainly concerned with here are Calcium, Magnesium, Potassium and Sodium. These are all alkaline cations, also called basic cations or bases. Both types of cations may be adsorbed onto either a clay particle or soil organic matter (SOM). All of the nutrients in the soil need to be held there somehow, or they will just wash away when you water the garden or get a good rainstorm. Clay particles almost always have a negative (-) charge, so they attract and hold positively (+) charged nutrients and non-nutrients. Soil organic matter (SOM) has both positive and negative charges, so it can hold on to both cations and anions.( http://www.soilminerals.com/Cation_Exchange_Simplified.htm)
Anion-a negatively charged ion (NO3–, PO42-, SO42-, etc…)
Soil particles and organic matter have negative charges on their surfaces. Mineral cations can adsorb to the negative surface charges or the inorganic and organic soil particles. Once adsorbed, these minerals are not easily lost when the soil is leached by water and they also provide a nutrient reserve available to plant roots.
These minerals can then be replaced or exchanged by other cations (i.e., cation exchange)
vThe exchage processes (Figure 23) are REVERSIBLE (unless something precipitates, volatilizes, or is strongly adsorbed).
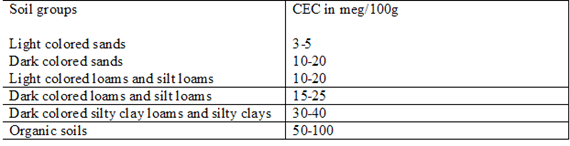
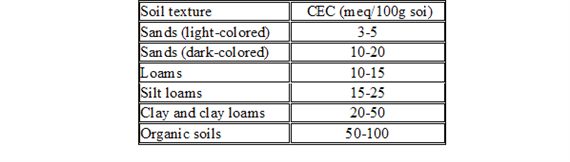
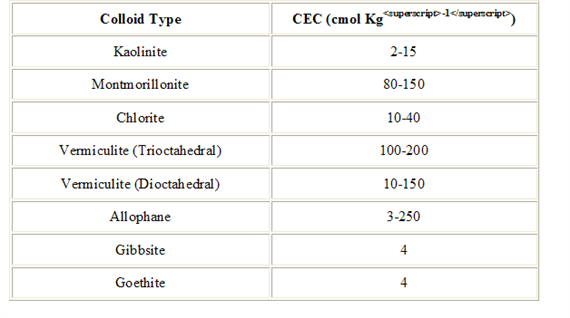
CEC is highly dependent upon soil texture and organic matter content Table 3, 4.). In general, the more clay and organic matter in the soil, the higher the CEC. Clay content is important because these small particles have a high ration of surface area to volume. Different types of clays also vary in CEC. Smectites have the highest CEC (80-100 millequivalents 100 g-1), followed by illites (15-40 meq 100 g-1) and kaolinites (3-15 meq 100 g-1).
Measurement of CEC.
The CEC of soil is usually measured by saturating the soil with an index cation such as Na+, removal of the excess salts of the index cation with a dilute solution, and then displacing the Na+ with another cation. The amount of Na+ displaced is then measured and the CEC is calculated.
In general, the CEC of most soils increases with an increase in soil pH. Two factors determine the relative proportions of the different cations adsorbed by clays. First, cations are not held equally tight by the soil colloids. When the cations are present in equivalent amounts, the order of strength of adsorption is Al3+ > Ca2+ > Mg2+ > K+ = NH4+ > Na+.
The relative concentrations of the cations in soil solution helps determine the degree of adsorption. Very acid soils will have high concentrations of H+ and Al3+. In neutral to moderately alkaline soils, Ca2+ and Mg2+ dominate. Poorly drained arid soils may adsorb Na in very high quantities.
Base saturation
The proportion of CEC satisfied by basic cations (Ca, Mg, K, and Na) is termed percentage base saturation (BS%). This property is inversely related to soil acidity. As the BS% increases, the pH increases. High base saturation is preferred but not essential for tree fruit production. The availability of nutrient cations such as Ca, Mg, and K to plants increases with increasing BS%.
Base saturation is usually close to 100% in arid region soils. Base saturation below 100% indicates that part of the CEC is occupied by hydrogen and/or aluminum ions. Base saturation above 100% indicates that soluble salts or lime may be present, or that there is a procedural problem with the analysis.
CEC and availability of nutrients
Exchangeable cations, may become available to plants. Plant roots also possess cation exchange capacity. Hydrogen ions from the root hairs and microorganisms may replace nutrient cations from the exchange complex on soil colloids. The nutrient cations are then released into the soil solution where they can be taken up by the adsorptive surfaces of roots and soil organisms. They may however, be lost from the system by drainage water.
Additionally, high levels of one nutrient may influence uptake of another (antagonistic relationship). For example, K uptake by plants is limited by high levels of Ca in some soils. High levels of K can in turn, limit Mg uptake even if Mg levels in soil are high.
Anion-exchange capacity (AEC)
Sources of anion exchange capacity
Anion exchange arise from the protonation of hydroxyl groups on the edges of silicate clays and on the surfaces of metal oxide clays Anion exchange is inversely related with pH is greatest in soils dominated by the sesquioxides. The anions Cl–, NO3–, and SeO42- and to some extent HS– ands SO42-, HCO3–, and CO3– adsorb mainly by ion exchange. Borate, phospahate and carboxylate adsorb principally by specific adsorption mechanisms. (http://jan.ucc.nau.edu/~doetqp-p/courses/env320/lec13/Lec13.html)
The total exchangeable anions that a soil can adsorb, measured as milliequivalents per 100 grams of soil. (http://www.encyclopedia.com/doc/1O7-anionexchangecapacity.html )
In contrast to CEC, AEC is the degree to which a soil can adsorb and exchange anions. AEC increases as soil pH decreases. The pH of most productive soils is usually too high (exceptions are for volcanic soils) for full development of AEC and thus it generally plays a minor role in supplying plants with anions.
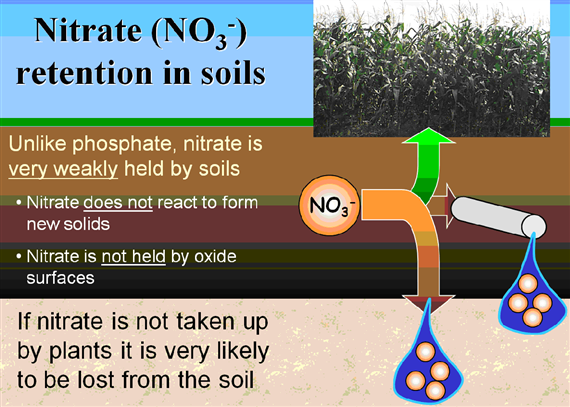
Because the AEC of most agricultural soils is small compared to their CEC, mineral anions such as nitrate (NO3– and Cl–) are repelled by the negative charge on soil colloids. These ions remain mobile in the soil solution and thus are susceptible to leaching. (http://soils.tfrec.wsu.edu/webnutritiongood/soilprops/04CEC.htm)
Phosphate anions are relativelly bounded on the positivelly charged places (iron, aluminium, calcium compounds etc.) (Figure 28).
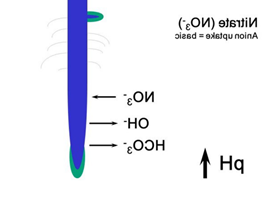
Nitrate is weakly bounded and the nitrate compounds are well soluble soluble in water, that why nitrate can easily be leahed out (Figure 29).
Nitrification
NH4+ NO3– Some of the ammonium produced by decomposition is converted to nitrate via a process called nitrification. The bacteria that carry out this reaction gain energy from it. Nitrification requires the presence of oxygen, so nitrification can happen only in oxygen-rich environments like circulating or flowing waters and the very surface layers of soils and sediments. The process of nitrification has some important consequences. Ammonium ions are positively charged and therefore stick (are sorbed) to negatively charged clay particles and soil organic matter. The positive charge prevents ammonium nitrogen from being washed out of the soil (or leached) by rainfall. In contrast, the negatively charged nitrate ion is not held by soil particles and so can be washed down the soil profile, leading to decreased soil fertility and nitrate enrichment of downstream surface and groundwaters.
Denitrification
NO3– N2+ N2O Through denitrification, oxidized forms of nitrogen such as nitrate and nitrite (NO2–) are converted to dinitrogen (N2) and, to a lesser extent, nitrous oxide gas. Denitrification is an anaerobic process that is carried out by denitrifying bacteria, which convert nitrate to dinitrogen in the following sequence:
NO3– NO2– NO N2O N2.
Nitric oxide and nitrous oxide are both environmentally important gases. Nitric oxide (NO) contributes to smog, and nitrous oxide (N2O) is an important greenhouse gas, thereby contributing to global climate change.
Once converted to dinitrogen, nitrogen is unlikely to be reconverted to a biologically available form because it is a gas and is rapidly lost to the atmosphere. Denitrification is the only nitrogen transformation that removes nitrogen from ecosystems (essentially irreversibly), and it roughly balances the amount of nitrogen fixed by the nitrogen fixers described above.
Denitrification of Nitrate-N
Certain soil bacteria that thrive in saturated (anaerobic) soil conditions will convert nitrate-N to oxygen and nitrogen gases. Volatilization of the nitrogen gas can result in N losses of as much as 5% of the available nitrate-N per day. Soils at greatest risk to denitrification N loss are those that are naturally heavy and poorly drained, plus fields with significant levels of soil compaction that restricts natural drainage. Because denitrification affects nitrate-N, the relative risk of N fertilizer products is identical to that for leaching N loss (Fig.16.).
Nitrogen immobilization
A fourth N loss mechanism is more temporary in nature. Soil microbes that decompose high carbon-content plant residues to organic matter use soil N during the decomposition process. Consequently, the nitrogen from the surface-applied fertilizer is “tied up” in the resulting organic matter and is temporarily unavailable for plant uptake until mineralization of the organic matter occurs at a later date. Such immobilization of soil N can be especially prevalent in high-residue no-till cropping systems. Unfortunately, applying N fertilizer in the fall to corn residues has not been shown to reduce N immobilization or speed residue decomposition. (http://www.agry.purdue.edu/ext/pubs/2006NLossMechanisms.pdf)
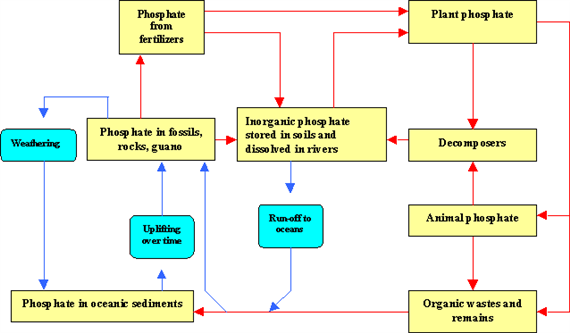
Phosphorus cycle
Phosphorus is an essential nutrient for plants and animals in the form of ions PO43- and HPO42-. It is a part of DNA-molecules, of molecules that store energy (ATP and ADP) and of fats of cell membranes. Phosphorus is also a building block of certain parts of the human and animal body, such as the bones and teeth.
Phosphorus can be found on earth in water, soil and sediments. Unlike the compounds of other matter cycles phosphorus cannot be found in air in the gaseous state. This is because phosphorus is usually liquid at normal temperatures and pressures. It is mainly cycling through water, soil and sediments. In the atmosphere phosphorus can mainly be found as very small dust particles.
Phosphorus moves slowly from deposits on land and in sediments, to living organisms, and than much more slowly back into the soil and water sediment. The phosphorus cycle is the slowest one of the matter cycles that are described here.
Phosphorus is most commonly found in rock formations and ocean sediments as phosphate salts. Phosphate salts that are released from rocks through weathering usually dissolve in soil water and will be absorbed by plants. Because the quantities of phosphorus in soil are generally small, it is often the limiting factor for plant growth. That is why humans often apply phosphate fertilizers on farmland. Phosphates are also limiting factors for plant-growth in marine ecosystems, because they are not very water-soluble. Animals absorb phosphates by eating plants or plant-eating animals.
Phosphorus cycles through plants and animals much faster than it does through rocks and sediments. When animals and plants die, phosphates will return to the soils or oceans again during decay. After that, phosphorus will end up in sediments or rock formations again, remaining there for millions of years. Eventually, phosphorus is released again through weathering and the cycle starts over (Fig. 17).
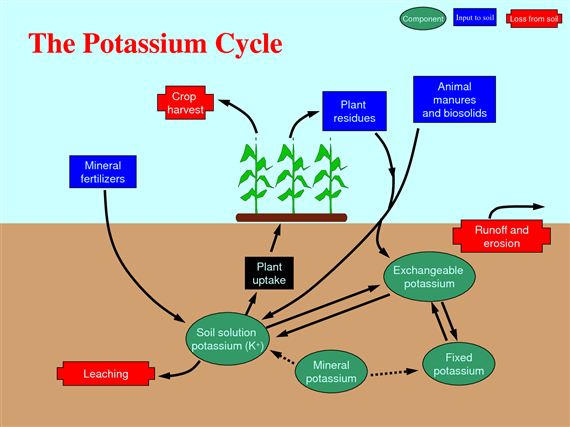
Potassium cycle
Potassium is taken up by plants in large quantities and is necessary to many plant functions, including carbohydrates metabolism, enzyme activation, osmotic regulation, and protein synthesis. Potassium is essential for photosynthesis, for nitrogen fixation in legumes, starch formation, and translocation of sugars. As a result of several of these functions, a good supply of potassium promotes production of plump grains and large tubers.
Potassium is important in helping plants adapt to environmental stresses (e.g. improved drought tolerance and winter hardiness, better resistance to fungal diseases and insect pests (Fig. 64).
Eutrophication
It means the gradual increase in the concentration of phosphorus, nitrogen, and other plant nutrients in an aging aquatic ecosystem such as a lake. The productivity or fertility of such an ecosystem increases as the amount of organic material that can be broken down into nutrients increases. This material enters the ecosystem primarily by runoff from land that carries debris and products of the reproduction and death of terrestrial organisms. Blooms, or great concentrations of algae and microscopic organisms, often develop on the surface, preventing the light penetration and oxygen absorption necessary for underwater life.
Figure 2.10 Cation uptake can have an acidic effect
Figure 2.12 Anion uptake can have a basic effect
Figure 2.11 Co-absorption of cations and anions at the same rate does not lead to a change in pH.
UF-AG Extension
—————————————————————
INTRODUCTION Iron is an essential nutrient for plants and serves as a cofactor for a wide variety of cellular processes, such as oxygen transport, cellular respiration, chlorophyll biosynthesis, thylakoid biogenesis and chloroplast development (Kobayashi and Nishizawa, 2012; Tanuja Poonia et al, 2018). The availability of Fe is severely limited in calcareous soils due to their low solubility at high pH and bicarbonate concentration which reduces the Fe uptake by inactivating the Fe in plants (Mortvedt, 1991; Najafi-Ghiri et al., 2013). Hence, Fe-deficiency induced chlorosis is a serious problem resulted in the yield loss and quality of crop produces in many crops particularly in the calcareous soils (Kim and Guerinot, 2007; Zheng, 2010). It is also closely related to the prevalence of Fe-deficiency-induced anemia in human beings (Murgia et al., 2012). Amelioration of Fe deficiency in soils and plants was generally achieved through the use of inorganic Fe salts, Fe chelates, organic manures, etc. either through soil application or as a foliar spray which differs significantly in maintaining soil Fe availability. Addition of Fe chelates to calcareous soils was proved to be very effective in maintaining soil solution Fe and the efficacy was better with Fe- EDDHA, Fe EDTA and Fe-DTPA but the recovery of Fe from ferrous sulphate was negligible under high soil pH and calcareousness (Jaloud et al., 2013; Faraz et al., 2014; Sedigheh Safarzadeh et al., 2018). Inclusion of organic manures proved to be beneficial in increasing the availability of Fe in soils and it was widely reported by many researchers (Ali et al., 2007; Yunchen Zhao, 2009; Amin, 2018). Further the Fe availability significantly correlated with many soil properties particularly pH, carbonate and bicarbonate ions and organic carbon content which majorly controls the availability (Obrador et al., 2007; Wang et al., 2009; Wu et al., 2010; Canasveras et al., 2014; Mahendra Kumar et al., 2017). Hence the present study was taken up to test the effectiveness of various levels and sources of Fe on Fe availability in calcareous and noncalcareous soils with and without amendments.
…
CONCLUSION To conclude, a linear increase in DTPA Fe extractability was observed with incubation period in all three soils and the highest availability was associated with the addition of 10 kg Fe EDTA ha-1 followed by 50 kg FeSO4 ha-1. Inclusion of FYM at 12.5 t ha-1 and 0.25 % Acetic acid considerably improved the Fe availability in soils and the better effect was registered with FYM. Higher Fe extractability was noted up to 30 days in the red calcareous soil while in black calcareous soils the release was linear up to 45 days. A negative correlation between soil pH and calcareousness was observed on the Fe availability in the soil
Calcareous soils (containing free lime) are common in many arid and semi-arid re gions af North America and occur as inclusions in mare humid regions. Phosphorus
(P) is very reactive with lime. Fallowing fertilizer application, P undergoes a series af reactions that gradually reduce its solubility. In mast calcareous sails, there does not appear ta be a strong agronomic advantage af any particular P source when managed properly. Organic matter can inhibit P fixation reactions ta same extent. SOILS fertilizer recommendations call far additional P ta be added when the sail contains high amounts af free lime. a lc ar eous soils are common in arid and semi-arid climates and occur as inclusions in more humid regions, af fecting over 1.5 billion acres of soil world wide and comprising more than 17% of the soils in the U.S. Calcareous soils are identi fied by the presence of the mineral calcium carbonate (CaC0 3 or lime) in the parent material and an accumulation of lime. This is most easily recognized by the efferves
cence (fizzing) that occurs when these soils are treated with dilute acid. The pH of these soils is usually above 7 and may be as high as 8.5. When these soils contain sodium carbonate, the pH may exceed 9. In some soils, CaC0 3 can concentrate into very hard layers, termed caliche, that are impermeable to water and plant roots. Calcareous soils can be extremely pro
surfaces, and pre cipit a ti onof various calcium phosphate min erals. While the total lime con tent of a soil is important for predicting P re actions, the lime particle size (and its effect on reac tive surface area) is often a better predictor of P behavior. Al though a calcar eous soil may be dominated by Effervescence (fizzing) occurs when colcoreous soils ore treoted with dilute ocid. Regulor soil testing is importont to monitor ovoilobility of P in colcoreous soils. ductive for agricultural use when they are managed properly. Since they are most fre quently found in semi-arid and arid re gions, supplemental irrigation water is of ten the frrst barrier for crop production. Limited availability of Pis often the next most limiting factor for plant growth. When P fertilizer is added to calcare ous soils, a series of fixation reactions oc cur that gradually decrease its solubility and eventually its availability to plants. Phosphorus “fixation” is a combination of surface adsorption on both clay and lime free lime, it may also contain significant amounts of iron (Fe), aluminum (Al), and manganese (Mn)… either as discrete min erals, as coatings on soil particles, or complexed with soil organic matter. These metals provide strong sorption sites for P and are frequently more significant in con trolling P solubility in calcareous soils than lime itself. Their importance should not be ignored. As fertilizer P reacts in calcareous soils, it is converted to less soluble compounds such as dicalcium phosphate dihydrate or
Figure 1. Fertilizer P undergoes a reduction in solubility following addition to three calcareous soils (Sharpley et al., 1989).
octacalcium phosphate. In some cases it may eventually convert to hydroxyapatite. A variety of management practices can he used to slow these natural fixation pro cesses and increase the efficiency of applied fertilizer for crop growth. A number of the factors controlling P availability will he briefly covered. Time—ln soluhle rock Pis treated af ter mining from geologic deposits to en hance its solubility and usefulness for plants. Fertilizer P is most soluble imme diately after addition to soil, then it un dergoes many chemical reactions that re sult in gradually diminished solubility (Figure 1).
Residual fertilizer P continues to he available for plant uptake for many years, but freshly applied P is generally most soluble and available for plant uptake. The common practice of building soil P concentrations to appropriate agronomic ranges provides a long-term source of this nutrient to crops. Phosphorus Fertilizer Source—Many studies have demonstrated that there are no consistent agronomic differences in most commercially available P fertilizers added to calcareous soils. The selection of a specific P source should he based on other factors such as application equipment, suitability of fluids or granules, and price. However, considerable work is cur rently underway to improve P availability with new P products and fertilizer addi tives. This topic will he explored in greater detail in future articles. For example, re cent work from Australia in extremely cal careous soils has suggested that fluid P sources may have somewhat greater solu bility and enhanced plant availability than granular fertilizers. It has been hypoth esized that granule dissolution may he sup pressed in these soil conditions. Additional work is underway in the U.S. to see if these results hold for soil conditions more typi cal of North America There is large variability in the solu bility and availability of P from various materials added to calcareous soil (Figure 2). These large differences are largely due to the unique properties of the materials, rather than any unique character associ ated with a specific soil. For example, the polymer-coated, slow release P source has very low apparent solubility, but is able to sup port high levels of plant P ac cumula tio n. The soluble P sources and liquid manures have very high solubility and also are able to maintain high P recovery by barley. Organic Matt e1′- l n the soil solution, there are several chemical components that will delay or prevent the reaction of P with lime. Organic mat ter has been found to interfere in the fixation reactions of P with lime. This inhibition of P
Figure 2. Extractability and P uptake by barley from various sources following incubation in a 12% lime soil. Sources initially added at a rate of 60 mg P/kg; extractions are average of 2 and 6-weeks sampling dates. (Leytem and Westerman, 2005).
Figure 3. The effect of soil temperature on fertilizer P extractability in a calcareous soil µavid and Rowell, 2003).
fixation may account for the observation that P availability is frequently greater in manured soils and with the addition of humic substances in lime-rich soil. Higher levels of soluble Fe, Al, and Mn are also related to increased P fixation in calcare ous soils. Temperature Soil temperature has two opposing effects on soil P availability. When fertilizer P is added to soil, it con tinually reacts and forms increasingly stable compounds for many months after application. The kinetics of the conversion of P to less soluble forms is more rapid under warmer conditions than incooler soil (Figure 3). An opposite effect occurs as increased soil temperature raises the solubility of soil P forms (both adsorbed or precipitated P). This well-known phenomenon accounts for frequent crop responses from added P in cool soils in the spring. In addition to im proved solubility, higher soil temperature increases P diffusion to plant roots and en hances overall root activity and prolifera tion. When planting early in the season, or in high-residue conditions, cold soil tem peratures can induce an early-stage P de ficiency in many types of soil. A starter P fertilizer application may help overcome these limitations. Adj11sting for Calcareous Soil s—Since the presence of lime in soils can reduce P availability to crops, fertilizer recommen dations are frequently adjusted to account for this condition. For example, the Uni versity of Idaho recommendations for potat rtilization state that an additional 10 lb Pp/A needs to he applied for every 1% increase in soil lime (Figure 4).
Figure 4. University of Idaho P fertilizer recom mendations for potatoes grown in calcareous soil take into account the free lime content of the soil (Tindall and Stark, 1997).
Calcareous soils can he extremely productive when managed properly. Maintaining an adequate supply of plant available P is essential to profitable and sustainable crop production. Since a vari ety of soil reactions tend to decrease the plant-availability of added fertilizer P in calcareous soil, regular soil testing should he conducted to avoid crop loss due to plant nutrient deficiency. ‘!ffl
The Nature of Phosphorus in Calcareous Soils, By A.B. Leytem and R.L. Mikkelsen, pdf
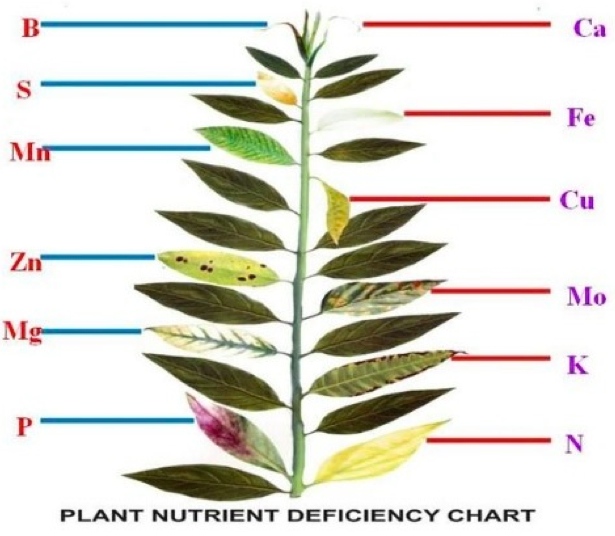
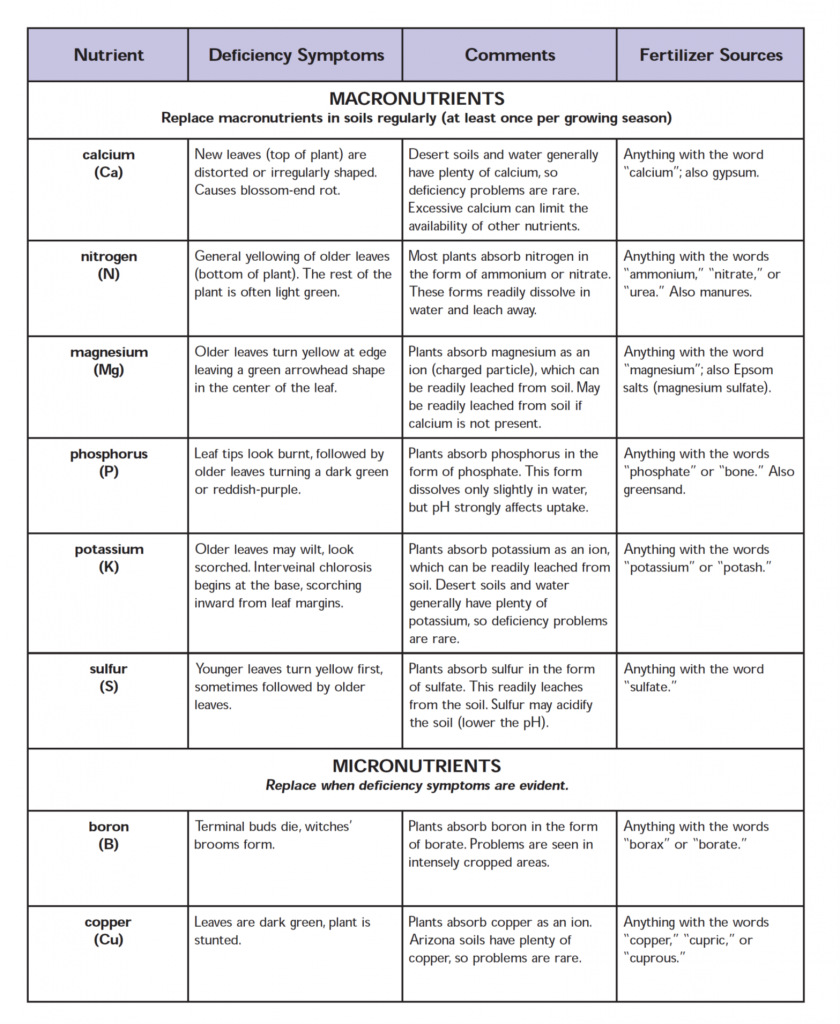
The most commonly found nutrient deficiency and toxicity symptoms are presented in the table below:
Nutrient
Deficiency Symptoms
Toxicity Symptoms
Nitrogen (N)
Stunted growth and restricted growth of lateral shoots. Plants express general chlorosis of the entire plant to light green and yellowing of older leaves which proceeds to younger leaves. Older leaves become necrotic and defoliate early
Plants are stunted, deep green in color, and secondary shoot development is poor. High N causes vegetative bud formation instead of reproductive bud formation. Ammonium toxicity can cause roots to turn brown, with necrotic root tips; reduce plant growth; necrotic lesions occur on stem and leaves; vascular browning occurs in stems and roots.
Phosphorus (P)
Stunted growth. Purplish coloration of older leaves in some plants. Dark green coloration with tips of leaves dying. Delayed maturity, Poor fruit and seed development.
Excess P in the plant can cause iron and zinc deficiencies.
Potassium (K)
Leaf margins turn chlorotic and then necrotic. Tip and marginal burn starting on mature leaves. Lower leaves turn yellow. Weak stalks and plant lodge easily. Slow growth.
High amounts of K can cause calcium (Ca), magnesium (Mg) and N deficiencies.
Magnesium (Mg)
Interveinal chlorosis on older leaves which proceeds to the younger leaves as the deficiency becomes more severe. The chlorotic interveinal yellow patches usually occur toward the center of the leaf with the margins being the last to turn yellow. Curling of leaves upward along margins.
High Mg can cause Ca deficiency.
Calcium (Ca)
Light green color on uneven chlorosis of young leaves. Brown or black scorching of new leaf tips and die-back of growing points. Growing points of stems and roots cease to develop. Poor root growth and roots short and thickened.
High Ca can cause Mg or Boron (B) deficiencies.
Sulfur (S)
Uniform chlorosis first appearing on new leaves.
Iron (Fe)
Interveinal chlorosis of new leaves followed by complete chlorosis and or bleaching of new leaves. Stunted growth.
Zinc (Zn)
Interveinal chlorosis of new leaves with some green next to veins. Short internodes and small leaves. Rosetting or whirling of leaves.
Manganese (Mn)
Interveinal chlorosis of new leaves with some green next to veins and later with grey or tan necrotic spots in chlorotic areas.
Copper (Cu)
Interveinal chlorosis of new leaves with tips and edges green, followed by veinal chlorosis. Leaves at the top of the plant wilt easily followed by chlorotic and necrotic areas in the leaves. Dieback of terminal shoots in trees.
Boron (B)
Death of terminal buds, causing lateral buds to develop and producing a ‘witches broom’ effect.
Symptoms develop as a yellow-tinted band around the leaf margins. The chlorotic zone becomes necrotic and gray, while the major portion of the leaf remains green.
Molybdenum (Mo)
Older leaves show interveinal chlorotic blotches, become cupped and thickened. Chlorosis continues upward to younger leaves as deficiency progresses.
Small, thick, and dark colored leaves. Lower leaves may turn yellow and brown, especially on the leaf margins. Plants become stunted. Roots are less vigorous, may have brown leaves, and are likely to become diseased.
The solution to high salt levels is usually to “leach out” (apply excess water to wash through the container) the growing medium or potting mix (termed the “root substrate” throughout this book).
Low EC ( see specific nutrient deficiency)
“chlorosis” (yellowing, resulting from lack of the green chlorophyll pigment that is essential for photosynthesis). These symptoms can be seen on this geraniu Nitrogen, phosphorus, and other nutrients are being moved out of its lower leaves to provide N and P to the growing points. The plant is essentially cannibalizing itself, because it cannot get nutrients from the roots. Growth and flowering can be greatly reduced
The solution to low fertilizer levels is simply to apply more fertilizer. However, it is important to diagnose that nutrient levels are indeed low using an onsite soil test method and checking EC is below the adequate range
Substrate pH is too high (alkaline, pH values above 7 are “basic”)
e.g. Iron Symptoms to look for with iron deficiency at high pH: Chlorosis either in the entire leaf or between the darker leaf veins (“interveinal chlorosis”) Symptoms show up first in young leaves, in contrast to an overall low nutrient level, which may be in all or older leaves. With sensitive plants such as the calibrachoa above, growth is severely stunted and growing points turn white and even necrotic (the tissue dies).
Reduce excess limestone, or alkaline water, nitrate fertilizer’s basic effects. To correct a high pH problem, a combination of acid fertilizer (containing ammonium or urea nitrogen) and adding acid into the irrigation water to remove alkalinity can usually drop substrate-pH. Drenches (irrigations applied to the root substrate) with additional iron in a highly soluble form are also very effective at corr
Substrate pH is too low (acidity)
Mn and Fe toxicity (over availability) Chlorotic (yellow) or necrotic (brown and dead) spots or leaf margins in older leaves, In marigold, the symptoms appear as brown sandy-colored spots in older leaves
The most common causes of a drop in substratepH are insufficient limestone, a fertilizer high in ammonium or urea nitrogen, which has an acidic reaction. Some plant species such as geranium also tend to drop substrate-pH. An application of a liming material such as flowable limestone or potassium bicarbonate, and a change to a basic nitrate fertilizer are needed to raise substrate-pH
ESSENTIAL NUTRIENTS, DEFICIENCIES, AND PLANT DISORDERS
Only certain elements have been determined to be essential for plant growth. An essential element is defined as one whose absence prevents a plant from completing its life cycle (Arnon and Stout 1939) or one that has a clear physiological role (Epstein 1999). If plants are given these essential elements, as well as energy from sunlight, they can synthesize all the compounds they need for normal growth. Table 5.1 lists the elements that are considered to be essential for most, if not all, higher plants. The first three elements—hydrogen, carbon, and oxygen—are not considered mineral nutrients because they are obtained primarily from water or carbon dioxide.
Essential mineral elements are usually classified as macronutrients or micronutrients, according to their relative concentration in plant tissue. In some cases, the differences in tissue content of macronutrients and micronutrients are not as great as those indicated in Table 5.1. For example, some plant tissues, such as the leaf mesophyll, have almost as much iron or manganese as they do sulfur or magnesium. Many elements often are present in concentrations greater than the plant’s minimum requirements.
Some researchers have argued that a classification into macronutrients and micronutrients is difficult to justify physiologically. Mengel and Kirkby (1987) have proposed that the essential elements be classified instead according to their biochemical role and physiological function. Table 5.2 shows such a classification, in which plant nutrients have been divided into four basic groups:
The first group of essential elements forms the organic (carbon carbon) compounds of the plant. Plants assimilate these nutrients via biochemical reactions involving oxidation and reduction.
The second group is important in energy storage reactions or in maintaining structural integrity. Elements in this group are often present in plant tissues as phosphate, borate, and silicate esters in which the elemental group is bound to the hydroxyl group of an organic molecule (i.e., sugar–phosphate).
The third group is present in plant tissue as either free ions or ions bound to substances such as the pectic acids present in the plant cell wall. Of particular importance are their roles as enzyme cofactors and in the regulation of osmotic potentials.
The fourth group has important roles in reactions involving electron transfer.
Naturally occurring elements, other than those listed in Table 5.1, can also accumulate in plant tissues. For example, aluminum is not considered to be an essential element, but plants commonly contain from 0.1 to 500 ppm aluminum, and addition of low levels of aluminum to a nutrient solution may stimulate plant growth Many species in the genera Astragalus, Xylorhiza, and Stanleya accumulate selenium, although plants have not been shown to have a specific requirement for this element.
Cobalt is part of cobalamin (vitamin B12 and its derivatives), a component of several enzymes in nitrogen-fixing microorganisms. Thus cobalt deficiency blocks the development and function of nitrogen-fixing nodules. Nonetheless, plants that do not fix nitrogen, as well as nitrogen-fixing plants that are supplied with ammonium or nitrate, do not require cobalt. Crop plants normally contain only relatively small amounts of nonessential elements.
Special Techniques Are Used in Nutritional Studies
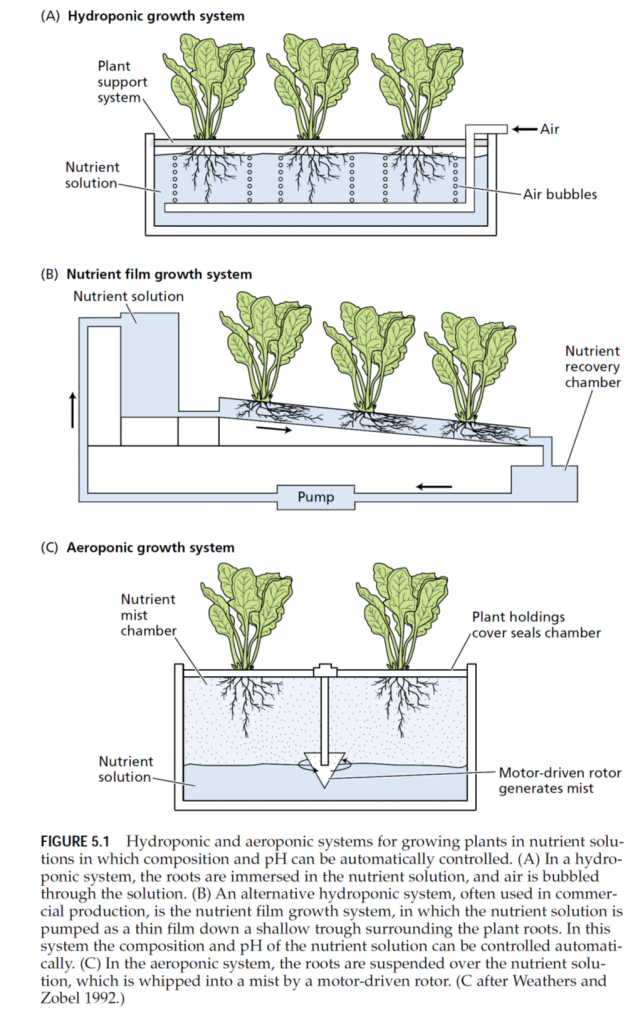
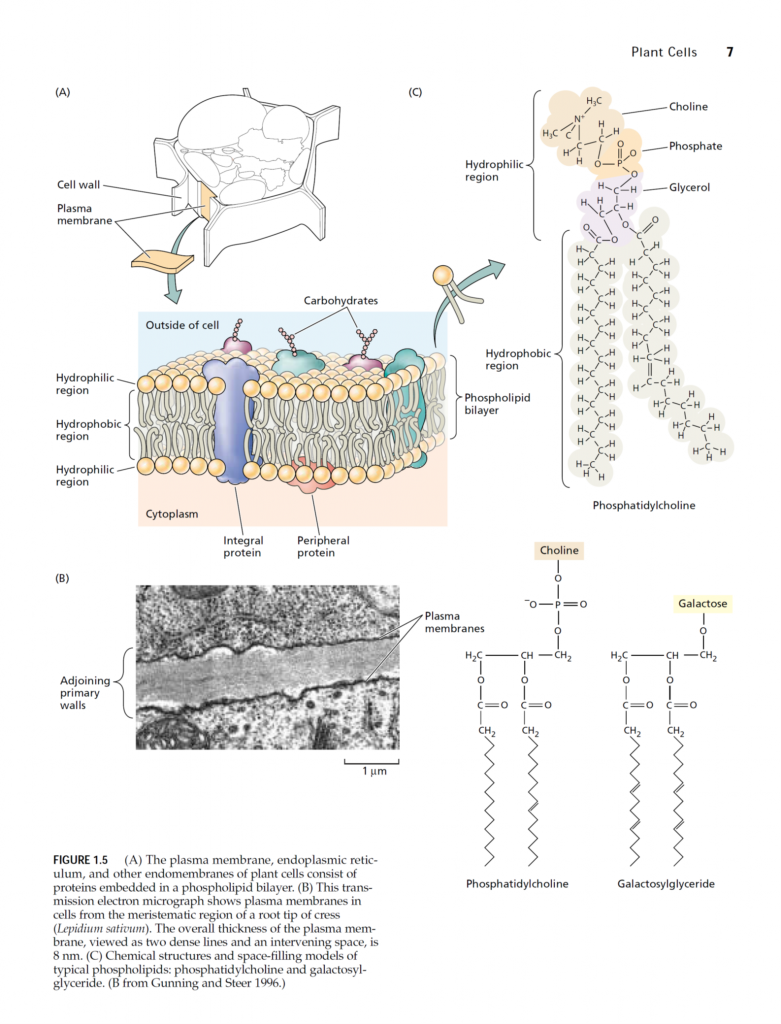
To demonstrate that an element is essential requires that plants be grown under experimental conditions in which only the element under investigation is absent. Such conditions are extremely difficult to achieve with plants grown in a complex medium such as soil. In the nineteenth century, several researchers, including Nicolas-Théodore de Saussure, Julius von Sachs, Jean-Baptiste-Joseph-Dieudonné Boussingault, and Wilhelm Knop, approached this problem by growing plants with their roots immersed in a nutrient solution containing only inorganic salts. Their demonstration that plants could grow normally with no soil or organic matter proved unequivocally that plants can fulfill all their needs from only inorganic elements and sunlight. The technique of growing plants with their roots immersed in nutrient solution without soil is called solution culture or hydroponics (Gericke 1937). Successful hydroponic culture (Figure 5.1A) requires a large volume of nutrient solution or frequent adjustment of the nutrient solution to prevent nutrient uptake by roots from producing radical changes in nutrient concentrations and pH of the medium. Asufficient supply of oxygen to the root system—also critical—may be achieved by vigorous bubbling of air through the medium. Hydroponics is used in the commercial production of many greenhouse crops. In one form of commercial hydroponic culture, plants are grown in a supporting material such as sand, gravel, vermiculite, or expanded clay (i.e., kitty litter). Nutrient solutions are then flushed through the supporting material, and old solutions are removed by leaching. In another form of hydroponic culture, plant roots lie on the surface of a trough, and nutrient solutions flow in a thin layer along the trough over the roots (Cooper 1979, Asher and Edwards 1983). This nutrient film growth system ensures that the roots receive an ample supply of oxygen (Figure 5.1B). Another alternative, which has sometimes been heralded as the medium of the future, is to grow the plants aeroponically (Weathers and Zobel 1992). In this technique, plants are grown with their roots suspended in air while being sprayed continuously with a nutrient solution (Figure 5.1C). This approach provides easy manipulation of the gaseous environment around the root, but it requires higher levels of nutrients than hydroponic culture does to sustain rapid plant growth. For this reason and other technical difficulties, the use of aeroponics is not widespread.
TABLE 5.1 Adequate tissue levels of elements that may be required by plants
Chemical Element symbol
Concentration in dry matter (% or ppm) a
Relative number of atoms with respect to molybdenum
Nutrient Solutions Can Sustain Rapid Plant Growth Over
the years, many formulations have been used for nutrient solutions. Early formulations developed by Knop in Germany included only KNO3, Ca(NO3)2, KH2PO4, MgSO4, and an iron salt. At the time this nutrient solution was believed to contain all the minerals required by the plant, but these experiments were carried out with chemicals that were contaminated with other elements that are now known to be essential (such as boron or molybdenum). Table 5.3 shows a more modern formulation for a nutrient solution. This formulation is called a modified Hoagland solution, named after Dennis R. Hoagland, a researcher who was prominent in the development of modern mineral nutrition research in the United States
A modified Hoagland solution contains all of the known mineral elements needed for rapid plant growth. The concentrations of these elements are set at the highest possible levels without producing toxicity symptoms or salinity stress and thus may be several orders of magnitude higher than those found in the soil around plant roots. For example, whereas phosphorus is present in the soil solution at concentrations normally less than 0.06 ppm, here it is offered at 62 ppm (Epstein 1972). Such high initial levels permit plants to be grown in a medium for extended periods without replenishment of the nutrients. Many researchers, however, dilute their nutrient solutions severalfold and replenish them frequently to minimize fluctuations of nutrient concentration in the medium and in plant tissue.
Another important property of the modified Hoagland formulation is that nitrogen is supplied as both ammonium (NH4 +) and nitrate (NO3 –). Supplying nitrogen in a balanced mixture of cations and anions tends to reduce the rapid rise in the pH of the medium that is commonly observed when the nitrogen is supplied solely as nitrate anion (Asher and Edwards 1983). Even when the pH of the medium is kept neutral, most plants grow better if they have access to both NH4 + and NO3 – because absorption and assimilation of the two nitrogen forms promotes cation–anion balance within the plant (Raven and Smith 1976; Bloom 1994).
A significant problem with nutrient solutions is maintaining the availability of iron. When supplied as an inorganic salt such as FeSO4 or Fe(NO3)2, iron can precipitate out of solution as iron hydroxide. If phosphate salts are present, insoluble iron phosphate will also form. Precipitation of the iron out of solution makes it physically unavailable to the plant, unless iron salts are added at frequent intervals. Earlier researchers approached this problem by adding iron together with citric acid or tartaric acid. Compounds such as these are called chelators because they form soluble complexes with cations such as iron and calcium in which the cation is held by ionic forces, rather than by covalent bonds. Chelated cations thus are physically more available to a plant. More modern nutrient solutions use the chemicals ethylenediaminetetraacetic acid (EDTA) or diethylenetriamine pentaacetic acid (DTPA, or pentetic acid) as chelating agents (Sievers and Bailar 1962). Figure 5.2 shows the structure of DTPA. The fate of the chelation complex during iron uptake by the root cells is not clear; iron may be released from the chelator when it is reduced from Fe3+ to Fe2+ at the root surface. The chelator may then diffuse back into the nutrient (or soil) solution and react with another Fe3+ ion or other metal ions. After uptake, iron is kept soluble by chelation with organic compounds present in plant cells. Citric acid may play a major role in iron chelation and its long-distance transport in the xylem.
Inadequate supply of an essential element results in a nutritional disorder manifested by characteristic deficiency symptoms. In hydroponic culture, withholding of an essential element can be readily correlated with a given set of symptoms for acute deficiencies. Diagnosis of soil-grown plants can be more complex, for the following reasons
Both chronic and acute deficiencies of several elements may occur simultaneously.
Deficiencies or excessive amounts of one element may induce deficiencies or excessive accumulations of another.
Some virus-induced plant diseases may producesymptoms similar to those of nutrient deficiencies.
Nutrient deficiency symptoms in a plant are the expression of metabolic disorders resulting from the insufficient supply of an essential element. These disorders are related to the roles played by essential elements in normal plant metabolism and function. Table 5.2 lists some of the roles of essential elements.
Even though each essential element participates in many different metabolic reactions, some general statements about the functions of essential elements in plant metabolism are possible. In general, the essential elements function in plant structure, metabolic function, and osmoregulation of plant cells. More specific roles may be related to the ability of divalent cations such as calcium or magnesium to modify the permeability of plant membranes. In addition, research continues to reveal specific roles of these elements in plant metabolism; for example, calcium acts as a signal to regulate key enzymes in the cytosol (Hepler and Wayne 1985; Sanders et al. 1999). Thus, most essential elements have multiple roles in plant metabolism.
When relating acute deficiency symptoms to a particular essential element, an important clue is the extent to which an element can be recycled from older to younger leaves. Some elements, such as nitrogen, phosphorus, and potassium, can readily move from leaf to leaf; others, such as boron, iron, and calcium, are relatively immobile in most plant species (Table 5.4). If an essential element is mobile, deficiency symptoms tend to appear first in older leaves. Deficiency of an immobile essential element will become evident first in younger leaves. Although the precise mechanisms of nutrient mobilization are not well understood, plant hormones such as cytokinins appear to be involved (see Chapter 21). In the discussion that follows, we will describe the specific deficiency symptoms and functional roles for the mineral essential elements as they are grouped in Table 5.2.
Group 1: Deficiencies in mineral nutrients that are part of carbon compounds.
This first group consists of nitro- gen and sulfur. Nitrogen availability in soils limits plant productivity in most natural and agricultural ecosystems. By contrast, soils generally contain sulfur in excess. Nonetheless, nitrogen and sulfur share the property that their oxidation–reduction states range widely (see Chapter 12). Some of the most energy-intensive reactions in life con- vert the highly oxidized, inorganic forms absorbed from the soil into the highly reduced forms found in organic compounds such as amino acids.
NITROGEN.
Nitrogen is the mineral element that plants require in greatest amounts. It serves as a constituent of many plant cell components, including amino acids and nucleic acids. Therefore, nitrogen deficiency rapidly inhibits plant growth. If such a deficiency persists, most species show chlorosis (yellowing of the leaves), especially in the older leaves near the base of the plant (for pictures of nitro- gen deficiency and the other mineral deficiencies described in this chapter, see Web Topic 5.1). Under severe nitrogen deficiency, these leaves become completely yellow (or tan) and fall off the plant. Younger leaves may not show these symptoms initially because nitrogen can be mobilized from older leaves. Thus a nitrogen-deficient plant may have light green upper leaves and yellow or tan lower leaves.
When nitrogen deficiency develops slowly, plants may have markedly slender and often woody stems. This wood- iness may be due to a buildup of excess carbohydrates that cannot be used in the synthesis of amino acids or other nitrogen compounds. Carbohydrates not used in nitrogen metabolism may also be used in anthocyanin synthesis, leading to accumulation of that pigment. This condition is revealed as a purple coloration in leaves, petioles, and stems of some nitrogen-deficient plants, such as tomato and certain varieties of corn.
SULFUR.
Sulfur is found in two amino acids and is a con- stituent of several coenzymes and vitamins essential for metabolism. Many of the symptoms of sulfur deficiency are similar to those of nitrogen deficiency, including chlorosis, stunting of growth, and anthocyanin accumulation. This similarity is not surprising, since sulfur and nitrogen are both constituents of proteins. However, the chlorosis caused by sulfur deficiency generally arises initially in mature and young leaves, rather than in the old leaves as in nitrogen deficiency, because unlike nitrogen, sulfur is not easily remobilized to the younger leaves in most species. Nonetheless, in many plant species sulfur chlorosis may occur simultaneously in all leaves or even initially in the older leaves.
Group 2: Deficiencies in mineral nutrients that are impor- tant in energy storage or structural integrity.
This group consists of phosphorus, silicon, and boron. Phosphorus and silicon are found at concentrations within plant tissue that warrant their classification as macronutrients, whereas boron is much less abundant and considered a micronutri- ent. These elements are usually present in plants as ester linkages to a carbon molecule.
Phosphorus (as phosphate, PO 3–) is an inte- gral component of important compounds of plant cells, including the sugar–phosphate intermediates of respiration and photosynthesis, and the phospholipids that make up plant membranes. It is also a component of nucleotides used in plant energy metabolism (such as ATP) and in DNA and RNA. Characteristic symptoms of phosphorus deficiency include stunted growth in young plants and a dark green coloration of the leaves, which may be mal- formed and contain small spots of dead tissue called necroticspots
As in nitrogen deficiency, some species may produce excess anthocyanins, giving the leaves a slight purple col- oration. In contrast to nitrogen deficiency, the purple col- oration of phosphorus deficiency is not associated with chlorosis. In fact, the leaves may be a dark greenish purple. Additional symptoms of phosphorus deficiency include the production of slender (but not woody) stems and the death of older leaves. Maturation of the plant may also be delayed.
SILICON.
Only members of the family Equisetaceae—called scouring rushes because at one time their ash, rich in gritty silica, was used to scour pots—require silicon to complete their life cycle. Nonetheless, many other species accumu- late substantial amounts of silicon within their tissues and show enhanced growth and fertility when supplied with adequate amounts of silicon (Epstein 1999).
Plants deficient in silicon are more susceptible to lodg- ing (falling over) and fungal infection. Silicon is deposited primarily in the endoplasmic reticulum, cell walls, and intercellular spaces as hydrated, amorphous silica (SiO2·nH2O). It also forms complexes with polyphenols and thus serves as an alternative to lignin in the reinforcement of cell walls. In addition, silicon can ameliorate the toxicity of many heavy metals.
BORON.
Although the precise function of boron in plant metabolism is unclear, evidence suggests that it plays roles in cell elongation, nucleic acid synthesis, hormone responses, and membrane function (Shelp 1993). Boron- deficient plants may exhibit a wide variety of symptoms, depending on the species and the age of the plant.
A characteristic symptom is black necrosis of the young leaves and terminal buds. The necrosis of the young leaves occurs primarily at the base of the leaf blade. Stems may be unusually stiff and brittle. Apical dominance may also be lost, causing the plant to become highly branched; how- ever, the terminal apices of the branches soon become necrotic because of inhibition of cell division. Structures such as the fruit, fleshy roots, and tubers may exhibit necro- sis or abnormalities related to the breakdown of internal tissues.
Group 3: Deficiencies in mineral nutrients that remain in ionic form.
This group includes some of the most familiar mineral elements: The macronutrients potassium, calcium, and magnesium, and the micronutrients chlorine, manganese, and sodium. They may be found in solution in the cytosol or vacuoles, or they may be bound electrostati- cally or as ligands to larger carbon-containing compounds.
Potassium, present within plants as the cation K+, plays an important role in regulation of the osmotic potential of plant cells (see Chapters 3 and 6). It also acti- vates many enzymes involved in respiration and photo- synthesis. The first observable symptom of potassium defi- ciency is mottled or marginal chlorosis, which then develops into necrosis primarily at the leaf tips, at the mar- gins, and between veins. In many monocots, these necrotic lesions may initially form at the leaf tips and margins and then extend toward the leaf base.
Because potassium can be mobilized to the younger leaves, these symptoms appear initially on the more mature leaves toward the base of the plant. The leaves may also curl and crinkle. The stems of potassium-deficient plants may be slender and weak, with abnormally short internodal regions. In potassium-deficient corn, the roots may have an increased susceptibility to root-rotting fungi present in the soil, and this susceptibility, together with effects on the stem, results in an increased tendency for the plant to be easily bent to the ground (lodging).
CALCIUM.
Calcium ions (Ca2+) are used in the synthesis of new cell walls, particularly the middle lamellae that sepa- rate newly divided cells. Calcium is also used in the mitotic spindle during cell division. It is required for the normal functioning of plant membranes and has been implicated as a second messenger for various plant responses to both environmental and hormonal signals (Sanders et al. 1999). In its function as a second messenger, calcium may bind to calmodulin, a protein found in the cytosol of plant cells. The calmodulin–calcium complex regulates many meta- bolic processes.
Characteristic symptoms of calcium deficiency include necrosis of young meristematic regions, such as the tips of roots or young leaves, where cell division and wall forma- tion are most rapid. Necrosis in slowly growing plants may be preceded by a general chlorosis and downward hook- ing of the young leaves. Young leaves may also appear deformed. The root system of a calcium-deficient plant may appear brownish, short, and highly branched. Severe stunting may result if the meristematic regions of the plant die prematurely.
MAGNESIUM.
In plant cells, magnesium ions (Mg2+) have a specific role in the activation of enzymes involved in respi- ration, photosynthesis, and the synthesis of DNA and RNA. Magnesium is also a part of the ring structure of the chloro- phyll molecule (see Figure 7.6A). A characteristic symptom of magnesium deficiency is chlorosis between the leaf veins, occurring first in the older leaves because of the mobility of this element. This pattern of chlorosis results because the chlorophyll in the vascular bundles remains unaffected for longer periods than the chlorophyll in the cells between the bundles does. If the deficiency is extensive, the leaves may become yellow or white. An additional symptom of mag- nesium deficiency may be premature leaf abscission.
CHLORINE.
The element chlorine is found in plants as the chloride ion (Cl–). It is required for the water-splitting reac- tion of photosynthesis through which oxygen is produced (see Chapter 7) (Clarke and Eaton-Rye 2000). In addition, chlorine may be required for cell division in both leaves and roots (Harling et al. 1997). Plants deficient in chlorine develop wilting of the leaf tips followed by general leaf chlorosis and necrosis. The leaves may also exhibit reduced growth. Eventually, the leaves may take on a bronzelike color (“bronzing”). Roots of chlorine-deficient plants may appear stunted and thickened near the root tips.
Chloride ions are very soluble and generally available in soils because seawater is swept into the air by wind and is delivered to soil when it rains. Therefore, chlorine defi- ciency is unknown in plants grown in native or agricultural habitats. Most plants generally absorb chlorine at levels much higher than those required for normal functioning.
MANGANESE.
Manganese ions (Mn2+) activate several enzymes in plant cells. In particular, decarboxylases and dehydrogenases involved in the tricarboxylic acid (Krebs) cycle are specifically activated by manganese. The best- defined function of manganese is in the photosynthetic reaction through which oxygen is produced from water (Marschner 1995). The major symptom of manganese defi- ciency is intervenous chlorosis associated with the devel- opment of small necrotic spots. This chlorosis may occur on younger or older leaves, depending on plant species and growth rate.
SODIUM.
Most species utilizing the C4 and CAM pathways of carbon fixation (see Chapter 8) require sodium ions (Na+). In these plants, sodium appears vital for regenerat- ing phosphoenolpyruvate, the substrate for the first car-boxylation in the C4 and CAM pathways (Johnstone et al. 1988). Under sodium deficiency, these plants exhibit chloro- sis and necrosis, or even fail to form flowers. Many C3 species also benefit from exposure to low levels of sodium ions. Sodium stimulates growth through enhanced cell expansion, and it can partly substitute for potassium as an osmotically active solute.
Group 4: Deficiencies in mineral nutrients that are involved in redox reactions.
This group of five micronu- trients includes the metals iron, zinc, copper, nickel, and molybdenum. All of these can undergo reversible oxidations and reductions (e.g., Fe2+ ~Fe3+) and have important roles in electron transfer and energy transformation. They are usu- ally found in association with larger molecules such as cytochromes, chlorophyll, and proteins (usually enzymes).
Iron has an important role as a component of enzymes involved in the transfer of electrons (redox reac- tions), such as cytochromes. In this role, it is reversibly oxi- dized from Fe2+ to Fe3+ during electron transfer. As in mag- nesium deficiency, a characteristic symptom of iron deficiency is intervenous chlorosis. In contrast to magne- sium deficiency symptoms, these symptoms appear ini- tially on the younger leaves because iron cannot be readily mobilized from older leaves. Under conditions of extreme or prolonged deficiency, the veins may also become chlorotic, causing the whole leaf to turn white.
The leaves become chlorotic because iron is required for the synthesis of some of the chlorophyll–protein complexes in the chloroplast. The low mobility of iron is probably due to its precipitation in the older leaves as insoluble oxides or phosphates or to the formation of complexes with phyto- ferritin, an iron-binding protein found in the leaf and other plant parts (Oh et al. 1996). The precipitation of iron dimin- ishes subsequent mobilization of the metal into the phloem for long-distance translocation.
ZINC.
Many enzymes require zinc ions (Zn2+) for their activity, and zinc may be required for chlorophyll biosyn- thesis in some plants. Zinc deficiency is characterized by a reduction in internodal growth, and as a result plants dis- play a rosette habit of growth in which the leaves form a circular cluster radiating at or close to the ground. The leaves may also be small and distorted, with leaf margins having a puckered appearance. These symptoms may result from loss of the capacity to produce sufficient amounts of the auxin indoleacetic acid. In some species (corn, sorghum, beans), the older leaves may become inter- venously chlorotic and then develop white necrotic spots. This chlorosis may be an expression of a zinc requirement for chlorophyll biosynthesis.
COPPER.
Like iron, copper is associated with enzymes involved in redox reactions being reversibly oxidized from Cu+ to Cu2+. An example of such an enzyme is plasto- cyanin, which is involved in electron transfer during the light reactions of photosynthesis (Haehnel 1984). The ini- tial symptom of copper deficiency is the production of dark green leaves, which may contain necrotic spots. The necrotic spots appear first at the tips of the young leaves and then extend toward the leaf base along the margins. The leaves may also be twisted or malformed. Under extreme copper deficiency, leaves may abscise prematurely.
NICKEL.
Urease is the only known nickel-containing enzyme in higher plants, although nitrogen-fixing microor- ganisms require nickel for the enzyme that reprocesses some of the hydrogen gas generated during fixation (hydrogen uptake hydrogenase) (see Chapter 12). Nickel- deficient plants accumulate urea in their leaves and, con- sequently, show leaf tip necrosis. Plants grown in soil sel- dom, if ever, show signs of nickel deficiency because the amounts of nickel required are minuscule.
MOLYBDENUM.
Molybdenum ions (Mo4+ through Mo6+) are components of several enzymes, including nitrate reductase and nitrogenase. Nitrate reductase catalyzes the reduction of nitrate to nitrite during its assimilation by the plant cell; nitrogenase converts nitrogen gas to ammonia in nitrogen-fixing microorganisms (see Chapter 12). The first indication of a molybdenum deficiency is general chloro- sis between veins and necrosis of the older leaves. In some plants, such as cauliflower or broccoli, the leaves may not become necrotic but instead may appear twisted and sub- sequently die (whiptail disease). Flower formation may be prevented, or the flowers may abscise prematurely.
Because molybdenum is involved with both nitrate assimilation and nitrogen fixation, a molybdenum defi- ciency may bring about a nitrogen deficiency if the nitrogen source is primarily nitrate or if the plant depends on sym- biotic nitrogen fixation. Although plants require only small amounts of molybdenum, some soils supply inadequate levels. Small additions of molybdenum to such soils can greatly enhance crop or forage growth at negligible cost.
TABLE 5.3 Composition of a modified Hoagland nutrient solution for growing plants
Compound
Molecular weight
Concentration of stock solution
Concentration of stock solution
Volume of stock solution per liter of final solution
Chelates Several factors reduce the bioavailability of Fe, including high soil pH, high bicarbonate content, plant species (grass species are usually more efficient than other species because they can excrete effective ligands), and abiotic stresses. Plants typically utilize iron as ferrous iron (Fe2+). Ferrous iron can be readily oxidized to the plant-unavailable ferric form (Fe3+) when soil pH is greater than 5.3 (Morgan and Lahav 2007). Iron deficiency often occurs if soil pH is greater than 7.4. Chelated iron can prevent this conversion from Fe2+ to Fe3+. Applying nutrients such as Fe, Mn, Zn, and Cu directly to the soil is inefficient because in soil solution they are present as positively charged metal ions and will readily react with oxygen and/or negatively charged hydroxide ions (OH-). If they react with oxygen or hydroxide ions, they form new compounds that are not bioavailable to plants. Both oxygen and hydroxide ions are abundant in soil and soilless growth media. The ligand can protect the micronutrient from oxidization or precipitation. Figure 1 shows examples of the typical iron deficiency symptoms of lychee grown in Homestead, Florida, in which the lychee trees have yellow leaves and small, abnormal fruits. Applying chelated fertilizers is an easy and practical correction method to avoid this nutrient disorder. For example, the oxidized form of iron is ferric (Fe3+), which is not bioavailable to plants and usually forms brown ferric hydroxide precipitation (Fe(OH)3). Ferrous sulfate, which is not a chelated fertilizer, is often used as the iron source. Its solution should be green. If the solution turns brown, the bioavailable form of iron has been oxidized and Fe is therefore unavailable to plants. Figure 3. Comparison of foliar applications of chelated Fe, regular iron fertilizers, and no iron fertilization for correcting iron deficiency of lychee(Litchi chinensis, the soapberry family). Credit: Yuncong Li, UF/IFAS [Pentetic acid or diethylenetriaminepentaacetic acid (DTPA)] The ligands EDTA, DTPA, and EDDHA are often used in chelated fertilizers (Table 4). Their effectiveness differs significantly. EDDHA chelated Fe is most stable at soil pH greater than 7 (Figure 4, A and B). Chelated fertilizer stability is desired because it means the chelated micronutrient will remain in a bioavailable form for a much longer time period, thus increasing micronutrient use efficiency in vegetable and fruit production. The stability of three typical chelated Fe fertilizers varies at different pH conditions (Figure 4, A). The Y-axis represents the ratio of chelated Fe to total chelate and ranges from 0 to 1.0. A value of 1.0 means the chelate is stable. The X-axis represents soil pH. At 6.0, the ratios for all three chelated Fe fertilizers are 1.0 (stable), but at pH 7.5, only the ratio of EDDTA chelated Fe is 1.0. That of DTPA chelated Fe is only 0.5, and that of EDTA chelated Fe is only 0.025. So, in practice, EDDTA chelated Fe fertilizer is most effective when pH is greater than 7 but most costly. Accordingly, crop yields of these three chelated fertilizers are in this order: FeEDDHA > FeDTPA > FeEDTA (Figure 4, B). See Micronutrient Deficiencies in Citrus: Iron, Zinc, and Manganese (https://edis.ifas.ufl.edu/ss423) for effective pH ranges of iron chelates. Table 3 shows the relationship between soil pH and chelated fertilizer requirement. Correction of Fe deficiency depends on individual crop response and many other factors. For instance, for vegetables, the rate is usually 0.4–1 lb. chelated Fe in 100 gal. of water per acre. Deciduous fruits need 0.1–0.2 lb. chelated Fe in 25 gal. of water per acre (Table 5). Foliar application is more effective than soil application. For foliar application, either inorganic or chelated Fe is effective, but for fertigation, chelated Fe should be used. In high pH soil, crops are also vulnerable to Cu deficiency stresses. Chelated Cu is significantly more effective than inorganic Cu. A commonly used copper chelate is Na2CuEDTA, which contains 13% Cu. Natural organic materials have approximately 0.5% Cu (Table 5). In addition to soil pH, Mn is also influenced by aeration, moisture, and organic matter content. Chelated Mn can improve Mn bioavailability. Mn deficiency occurs more often in high pH and dry soil. Similar to other micronutrients, foliar spray is much more effective than soil application. For commercial vegetable production, 0.2–0.5 lb. MnEDTA in 200 gal. of water per acre can effectively correct Mn deficiency (Table 5). Zinc is another micronutrient whose bioavailability is closely associated with soil pH. Crops may be susceptible to Zn deficiency in soil with pH > 7.3. Spraying 0.10–0.14 lb. chelated Zn in 100 gal. of water per acre is effective (Poh et al. 2009). Animal waste and municipal waste also contain Cu, Mn, and Zn micronutrients (Table 5). For more information about micronutrient deficiency in crops, see Plant Tissue Analysis and Interpretation for Vegetable Crops in Florida (https://edis.ifas.ufl.edu/ep081),
TABLE 5.5 Comparison of properties of three major types of silicate clays found in the soil
Analysis of Plant Tissues Reveals Mineral Deficiencies
Requirements for mineral elements change during the growth and development of a plant. In crop plants, nutri- ent levels at certain stages of growth influence the yield of the economically important tissues (tuber, grain, and so on). To optimize yields, farmers use analyses of nutrient levels in soil and in plant tissue to determine fertilizer schedules.
Soilanalysisis the chemical determination of the nutri- ent content in a soil sample from the root zone. As dis- cussed later in the chapter, both the chemistry and the biol- ogy of soils are complex, and the results of soil analyses vary with sampling methods, storage conditions for the samples, and nutrient extraction techniques. Perhaps more important is that a particular soil analysis reflects the lev- els of nutrients potentially available to the plant roots from the soil, but soil analysis does not tell us how much of a particular mineral nutrient the plant actually needs or is able to absorb. This additional information is best deter- mined by plant tissue analysis.
Proper use of plant tissue analysis requires an under- standing of the relationship between plant growth (or yield) and the mineral concentration of plant tissue sam- ples (Bouma 1983). As the data plot in Figure 5.3 shows, when the nutrient concentration in a tissue sample is low, growth is reduced. In this deficiency zone of the curve, an increase in nutrient availability is directly related to an increase in growth or yield. As the nutrient availability con- tinues to increase, a point is reached at which further addi- tion of nutrients is no longer related to increases in growth or yield but is reflected in increased tissue concentrations. This region of the curve is often called the adequate zone. The transition between the deficiency and adequate zones of the curve reveals the critical concentration of the nutrient (see Figure 5.3), which may be defined as the min- imum tissue content of the nutrient that is correlated with maximal growth or yield. As the nutrient concentration of the tissue increases beyond the adequate zone, growth or yield declines because of toxicity (this is the toxic zone).
To evaluate the relationship between growth and tissue nutrient concentration, researchers grow plants in soil or nutrient solution in which all the nutrients are present in adequate amounts except the nutrient under consideration. At the start of the experiment, the limiting nutrient is added in increasing concentrations to different sets of plants, and the concentrations of the nutrient in specific tis- sues are correlated with a particular measure of growth or yield. Several curves are established for each element, one for each tissue and tissue age.
Because agricultural soils are often limited in the ele- ments nitrogen, phosphorus, and potassium, many farm- ers routinely use, at a minimum, curves for these elements. If a nutrient deficiency is suspected, steps are taken to cor- rect the deficiency before it reduces growth or yield. Plant analysis has proven useful in establishing fertilizer sched- ules that sustain yields and ensure the food quality of many crops.
TREATING NUTRITIONAL DEFICIENCIES
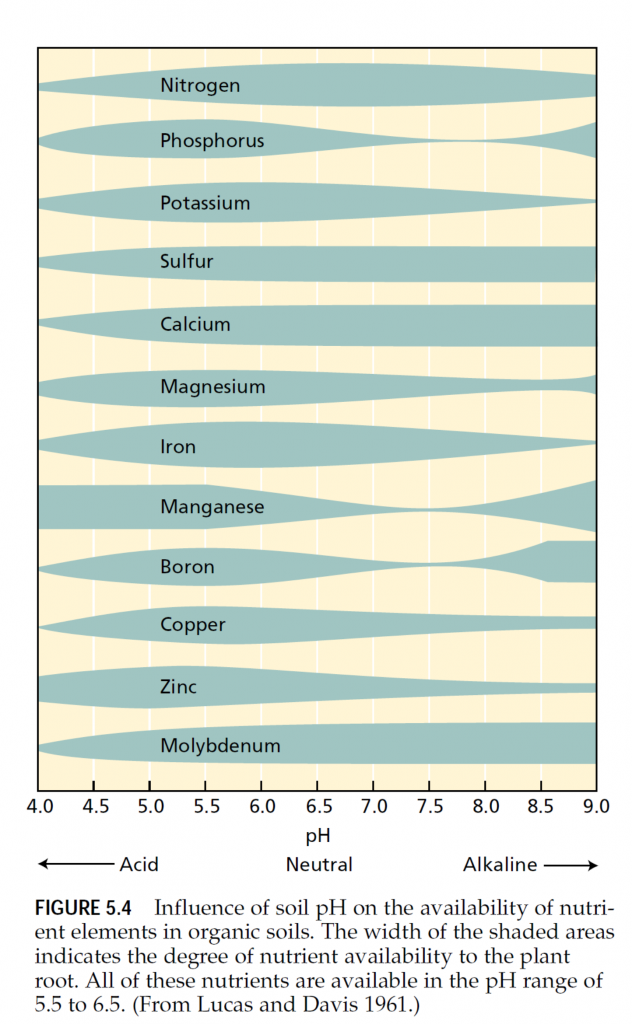
Many traditional and subsistence farming practices pro- mote the recycling of mineral elements. Crop plants absorb the nutrients from the soil, humans and animals consume locally grown crops, and crop residues and manure from humans and animals return the nutrients to the soil. The main losses of nutrients from such agricultural systems ensue from leaching that carries dissolved ions away with drainage water. In acid soils, leaching may be decreased by the addition of lime—a mix of CaO, CaCO3, and Ca(OH)2—to make the soil more alkaline because many mineral elements form less soluble compounds when the pH is higher than 6 (Figure 5.4).
In the high-production agricultural systems of industrial countries, the unidirectional removal of nutrients from the soil to the crop can become significant because a large por- tion of crop biomass leaves the area of cultivation. Plants synthesize all their components from basic inorganic sub- stances and sunlight, so it is important to restore these lost nutrients to the soil through the addition of fertilizers.
Crop Yields Can Be Improved by Addition of Fertilizers
Most chemical fertilizers contain inorganic salts of the macronutrients nitrogen, phosphorus, and potassium (see Table 5.1). Fertilizers that contain only one of these three nutrients are termed straightfertilizers.Some examples of straight fertilizers are superphosphate, ammonium nitrate, and muriate of potash (a source of potassium). Fertilizers that contain two or more mineral nutrients are called com- pound fertilizers or mixedfertilizers, and the numbers on the package label, such as 10-14-10, refer to the effective per- centages of N, P2O5, and K2O, respectively, in the fertilizer.
With long-term agricultural production, consumption of micronutrients can reach a point at which they, too, must be added to the soil as fertilizers. Adding micronutrients to the soil may also be necessary to correct a preexisting defi- ciency. For example, some soils in the United States are
water and oxygen availability, and the type and number of microorganisms present in the soil.
As a consequence, the rate of mineralization is highly variable, and nutrients from organic residues become avail- able to plants over periods that range from days to months to years. The slow rate of mineralization hinders efficient fertilizer use, so farms that rely solely on organic fertilizers may require the addition of substantially more nitrogen or phosphorus and suffer even higher nutrient losses than farms that use chemical fertilizers. Residues from organic fertilizers do improve the physical structure of most soils, enhancing water retention during drought and increasing drainage in wet weather.
TABLE 5.4 Mineral elements classified on the basis of their mobility within a plant and their tendency to retranslocate during deficiencies
Mobile
Immobile
Nitrogen
Calcium
Potassium
Sulfur
Magnesium
Iron
Phosphorus
Boron
Chlorine
Copper
Sodium
Zinc
Molybdenum
Some Mineral Nutrients Can Be Absorbed by Leaves
In addition to nutrients being added to the soil as fertiliz- ers, some mineral nutrients can be applied to the leaves as sprays, in a process known as foliar application, and the leaves can absorb the applied nutrients. In some cases, this method can have agronomic advantages over the applica- tion of nutrients to the soil. Foliar application can reduce the lag time between application and uptake by the plant, which could be important during a phase of rapid growth. It can also circumvent the problem of restricted uptake of a nutrient from the soil. For example, foliar application of mineral nutrients such as iron, manganese, and copper may be more efficient than application through the soil
deficient in boron, copper, zinc, manganese, molybdenum, or iron (Mengel and Kirkby 1987) and can benefit from nutrient supplementation.
Chemicals may also be applied to the soil to modify soil pH. As Figure 5.4 shows, soil pH affects the availability of all mineral nutrients. Addition of lime, as mentioned previ- ously, can raise the pH of acidic soils; addition of elemental sulfur can lower the pH of alkaline soils. In the latter case, microorganisms absorb the sulfur and subsequently release sulfate and hydrogen ions that acidify the soil.
Organic fertilizers, in contrast to chemical fertilizers, originate from the residues of plant or animal life or from natural rock deposits. Plant and animal residues contain many of the nutrient elements in the form of organic com- pounds. Before crop plants can acquire the nutrient ele- ments from these residues, the organic compounds must be broken down, usually by the action of soil microorgan- isms through a process called mineralization. Mineraliza- tion depends on many factors, including temperature, available to the root system.
Nutrient uptake by plant leaves is most effective when the nutrient solution remains on the leaf as a thin film (Mengel and Kirkby 1987). Production of a thin film often requires that the nutrient solutions be supplemented with surfactant chemicals, such as the detergent Tween 80, that reduce surface tension. Nutrient movement into the plant seems to involve diffusion through the cuticle and uptake by leaf cells. Although uptake through the stomatal pore could provide a pathway into the leaf, the architecture of the pore (see Figures 4.13 and 4.14) largely prevents liquid penetration (Ziegler 1987).
For foliar nutrient application to be successful, damage to the leaves must be minimized. If foliar sprays are applied on a hot day, when evaporation is high, salts may accumulate on the leaf surface and cause burning or scorching. Spraying on cool days or in the evening helps to alleviate this problem. Addition of lime to the spray dimin- ishes the solubility of many nutrients and limits toxicity. Foliar application has proved economically successful mainly with tree crops and vines such as grapes, but it is also used with cereals. Nutrients applied to the leaves could save an orchard or vineyard when soil-applied nutri- ents would be too slow to correct a deficiency. In wheat, nitrogen applied to the leaves during the later stages of growth enhances the protein content of seeds.
…
SUMMARY
Plants are autotrophic organisms capable of using the energy from sunlight to synthesize all their components from carbon dioxide, water, and mineral elements. Studies of plant nutrition have shown that specific mineral ele- ments are essential for plant life. These elements are clas- sified as macronutrients or micronutrients, depending on the relative amounts found in plant tissue.
Certain visual symptoms are diagnostic for deficiencies in specific nutrients in higher plants. Nutritional disorders occur because nutrients have key roles in plant metabolism. They serve as components of organic compounds, in energy storage, in plant structures, as enzyme cofactors, and in electron transfer reactions. Mineral nutrition can be studied through the use of hydroponics or aeroponics, which allow the characterization of specific nutrient requirements. Soil and plant tissue analysis can provide information on the nutritional status of the plant–soil sys- tem and can suggest corrective actions to avoid deficien- cies or toxicities.
When crop plants are grown under modern high-pro- duction conditions, substantial amounts of nutrients are removed from the soil. To prevent the development of defi- ciencies, nutrients can be added back to the soil in the form of fertilizers. Fertilizers that provide nutrients in inorganic forms are called chemical fertilizers; those that derive from plant or animal residues are considered organic fertilizers. In both cases, plants absorb the nutrients primarily as inor- ganic ions. Most fertilizers are applied to the soil, but some are sprayed on leaves.
Biological Membranes Are Phospholipid Bilayers That Contain Proteins
All cells are enclosed in a membrane that serves as their outer boundary, separating the cytoplasm from the exter- nal environment. This plasmamembrane(also called plas-malemma) allows the cell to take up and retain certain sub- stances while excluding others. Various transport proteins embedded in the plasma membrane are responsible for this selective traffic of solutes across the membrane. The accu- mulation of ions or molecules in the cytosol through the action of transport proteins consumes metabolic energy. Membranes also delimit the boundaries of the specialized internal organelles of the cell and regulate the fluxes of ions and metabolites into and out of these compartments.
According to the fluid-mosaic model, all biological membranes have the same basic molecular organization. They consist of a double layer (bilayer) of either phospho- lipids or, in the case of chloroplasts, glycosylglycerides, in which proteins are embedded (Figure 1.5A and B). In most membranes, proteins make up about half of the mem- brane’s mass. However, the composition of the lipid com- ponents and the properties of the proteins vary from mem- brane to membrane, conferring on each membrane its unique functional characteristics
Phospholipids. Phospholipids are a class of lipids in which two fatty acids are covalently linked to glycerol, which is covalently linked to a phosphate group. Also attached to this phosphate group is a variable component, called the head group, such as serine, choline, glycerol, or inositol (Figure 1.5C). In contrast to the fatty acids, the head groups are highly polar; consequently, phospholipid mol- ecules display both hydrophilic and hydrophobic proper- ties (i.e., they are amphipathic). The nonpolar hydrocarbon chains of the fatty acids form a region that is exclusively hydrophobic—that is, that excludes water.
Plastid membranes are unique in that their lipid com- ponent consists almost entirely of glycosylglyceridesrather than phospholipids. In glycosylglycerides, the polar head group consists of galactose, digalactose, or sulfated galactose, without a phosphate group. The fatty acid chains of phospholipids and glycosyl- glycerides are variable in length, but they usually consist of 14 to 24 carbons. One of the fatty acids is typically saturated (i.e., it contains no double bonds); the other fatty acid chain usually has one or more cis double bonds (i.e., it is unsaturated).
The presence of cis double bonds creates a kink in the chain that prevents tight packing of the phospholipids in the bilayer. As a result, the fluidity of the membrane is increased. The fluidity of the membrane, in turn, plays a critical role in many membrane functions. Membrane flu- idity is also strongly influenced by temperature. Because plants generally cannot regulate their body temperatures, they are often faced with the problem of maintaining mem- brane fluidity under conditions of low temperature, which tends to decrease membrane fluidity. Thus, plant phos- pholipids have a high percentage of unsaturated fatty acids, such as oleic acid (one double bond), linoleic acid (two double bonds) and a-linolenic acid (three double bonds), which increase the fluidity of their membranes.
Proteins.
The proteins associated with the lipid bilayer are of three types: integral, peripheral, and anchored. Integralproteinsare embedded in the lipid bilayer. Most inte- gral proteins span the entire width of the phospholipid bilayer, so one part of the protein interacts with the outside of the cell, another part interacts with the hydrophobic core of the membrane, and a third part interacts with the inte- rior of the cell, the cytosol. Proteins that serve as ion chan- nels (see Chapter 6) are always integral membrane pro- teins, as are certain receptors that participate in signal transduction pathways (see Chapter 14). Some receptor-like proteins on the outer surface of the plasma membrane rec- ognize and bind tightly to cell wall consituents, effectively cross-linking the membrane to the cell wall.
Peripheralproteinsare bound to the membrane surface by noncovalent bonds, such as ionic bonds or hydrogen bonds, and can be dissociated from the membrane with high salt solutions or chaotropic agents, which break ionic and hydrogen bonds, respectively. Peripheral proteins serve a variety of functions in the cell. For example, some are involved in interactions between the plasma membrane and components of the cytoskeleton, such as microtubules and actin microfilaments, which are discussed later in this chapter.
Anchored proteins are bound to the membrane surface via lipid molecules, to which they are covalently attached. These lipids include fatty acids (myristic acid and palmitic acid), prenyl groups derived from the isoprenoid pathway (farnesyl and geranylgeranyl groups), and glycosylphos- phatidylinositol (GPI)-anchored proteins (Figure 1.6) (Buchanan et al. 2000).
The Nucleus Contains Most of the Genetic Material of the Cell
The nucleus (plural nuclei) is the organelle that contains the genetic information primarily responsible for regulating the metabolism, growth, and differentiation of the cell. Collectively, these genes and their intervening sequences are referred to as the nuclear genome. The size of the nuclear genome in plants is highly variable, ranging from about 1.2 × 108 base pairs for the diminutive dicot Arabidopsis thaliana to 1 × 1011 base pairs for the lily Fritillaria assyriaca. The remainder of the genetic information of the cell is contained in the two semiautonomous organelles—the chloroplasts and mitochondria—which we will discuss a little later in this chapter. The nucleus is surrounded by a double membrane called the nuclear envelope (Figure 1.7A). The space between the two membranes of the nuclear envelope is called the perinuclear space, and the two membranes of the nuclear envelope join at sites called nuclear pores (Figure 1.7B).
Nuclear import and export through nuclear pore complexes. An import complex consisting of an NLS-bearing cargo and a nuclear transport receptor (NTR) is formed in the cytoplasm. After translocation through the NPC, Ran-GTP displaces the cargo from the NTR, resulting in nuclear cargo release. This reaction occurs due to the chromatin localization of the Ran guanine nucleotide exchange factor (RanGEF), which is restricted to the nucleus. The NTR–Ran-GTP complex returns to the cytoplasm through the NPC where the Ran GTPase-activating protein (RanGAP1) stimulates GTP hydrolysis, releasing the NTR for another import cycle. Nuclear export cycles require the formation of a trimeric cargo–NTR–Ran-GTP complex in the nucleus. After NPC passage, this complex dissociates due to Ran-GTP hydrolysis, releasing the cargo into the cytoplasm. Perforating the nuclear boundary – how nuclear pore complexes assemble
The nuclear “pore” is actually an elaborate structure composed of more than a hundred different proteins arranged octagonally to form a nuclear pore complex (Figure 1.8). There can be very few to many thousands of nuclear pore complexes on an individual nuclear envelope. The central “plug” of the complex acts as an active (ATPdriven) transporter that facilitates the movement of macromolecules and ribosomal subunits both into and out of the nucleus. (Active transport will be discussed in detail in Chapter 6.) A specific amino acid sequence called the nuclear localization signal is required for a protein to gain entry into the nucleus. The nucleus is the site of storage and replication of the chromosomes, composed of DNA and its associated proteins. Collectively, this DNA–protein complex is known chromatin. The linear length of all the DNA within any plant genome is usually millions of times greater than the diameter of the nucleus in which it is found. To solve the problem of packaging this chromosomal DNA within the nucleus, segments of the linear double helix of DNA are coiled twice around a solid cylinder of eight histone protein molecules, forming a nucleosome. Nucleosomes are arranged like beads on a string along the length of each chromosome. During mitosis, the chromatin condenses, first by coiling tightly into a 30 nm chromatin fiber, with six nucleosomes per turn, followed by further folding and packing processes that depend on interactions between proteins and nucleic acids (Figure 1.9). At interphase, two types of chromatin are visible: heterochromatin and euchromatin. About 10% of the DNA consists of heterochromatin, a highly compact and transcriptionally inactive form of chromatin. The rest of the DNA consists of euchromatin, the dispersed, transcriptionally active form. Only about 10% of the euchromatin is transcriptionally active at any given time. The remainder exists in an intermediate state of condensation, between heterochromatin and transcriptionally active euchromatin. Nuclei contain a densely granular region, called the nucleolus (plural nucleoli), that is the site of ribosome synthesis (see Figure 1.7A). The nucleolus includes portions of one or more chromosomes where ribosomal RNA (rRNA) genes are clustered to form a structure called the nucleolar organizer. Typical cells have one or more nucleoli per nucleus. Each 80S ribosome is made of a large and a small subunit, and each subunit is a complex aggregate of rRNA and specific proteins. The two subunits exit the nucleus separately, through the nuclear pore, and then unite in the cytoplasm to form a complete ribosome (Figure 1.10A). Ribosomes are the sites of protein synthesis. Protein Synthesis Involves Transcription and Translation The complex process of protein synthesis starts with transcription— the synthesis of an RNA polymer bearing a base Translation is the process whereby a specific protein is synthesized from amino acids, according to the sequence information encoded by the mRNA. The ribosome travels the entire length of the mRNA and serves as the site for the sequential bonding of amino acids as specified by the base sequence of the mRNA (Figure 1.10B).
Cells have an elaborate network of internal membranes called the endoplasmic reticulum (ER). The membranes of the ER are typical lipid bilayers with interspersed integral and peripheral proteins. These membranes form flattened or tubular sacs known as cisternae (singular cisterna). Ultrastructural studies have shown that the ER is continuous with the outer membrane of the nuclear envelope. There are two types of ER—smooth and rough (Figure 1.11)—and the two types are interconnected. Rough ER (RER) differs from smooth ER in that it is covered with ribosomes that are actively engaged in protein synthesis; in addition, rough ER tends to be lamellar (a flat sheet composed of two unit membranes), while smooth ER tends to be tubular, although a gradation for each type can be observed in almost any cell. The structural differences between the two forms of ER are accompanied by functional differences. Smooth ER functions as a major site of lipid synthesis and membrane assembly. Rough ER is the site of synthesis of membrane proteins and proteins to be secreted outside the cell or into the vacuoles.
Transport pathways between the ER and the Golgi complex. COPII vesicles exiting from the ER (shown on the left) carry transport machinery (black), membrane phospholipids (red), membrane cargo proteins (green), fluid (light blue) and captured soluble cargo (dark blue) in the forward direction, as indicated by the colored arrows (1). COPI vesicles (shown on the right) retrieve the machinery to the ER but might be insufficient to also recycle lipids and fluid. This concept is schematically illustrated by the relative lengths of the colored arrows representing COPII-dependent forward transport (1) and COPI-dependent retrograde transport (2). Fluid content of the ER could re-equilibrate with the cytosol (curved light-blue arrows at the bottom of the figure), whereas phospholipids could be returned by Rab6-dependent tubular carriers (arrows, 3). The imbalance of fluid transport between the anterograde and retrograde directions could be responsible for a valve-like system that ensures movement in the forward direction of soluble cargo (light and dark blue arrows, 4). Membrane cargo could be captured into the COPI-independent retrograde carriers more efficiently than fluid could, because of the high surface-to-volume ratio of tubules; this partitioning phenomenon is expected to cause recycling of membrane cargo (green arrow, 3) with a consequent delay in their anterograde transport. Membrane cargo that escapes this recycling phenomenon progresses further through the Golgi (green arrow, 4). Membrane cargo proteins that carry export signals are not represented in this cartoon. Getting membrane proteins on and off the shuttle bus between the endoplasmic reticulum and the Golgi complex
Secretion of Proteins from Cells Begins with the Rough ER
Proteins destined for secretion cross the RER membrane and enter the lumen of the ER. This is the first step in the secretion pathway that involves the Golgi body and vesicles that fuse with the plasma membrane. The mechanism of transport across the membrane is complex, involving the ribosomes, the mRNA that codes for the secretory protein, and a special receptor in the ER membrane. All secretory proteins and most integral membrane proteins have been shown to have a hydrophobic sequence of 18 to 30 amino acid residues at the amino-terminal end of the chain. During translation, this hydrophobic leader, called the signal peptide sequence, is recognized by a signal recognition particle (SRP), made up of protein and RNA, which facilitates binding of the free ribosome to SRP receptor proteins (or “docking proteins”) on the ER (see Figure 1.10A). The signal peptide then mediates the transfer of the elongating polypeptide across the ER membrane into the lumen. (In the case of integral membrane proteins, a portion of the completed polypeptide remains embedded in the membrane.) Once inside the lumen of the ER, the signal sequence is cleaved off by a signal peptidase. In some cases, a branched oligosaccharide chain made up of N-acetylglucosamine (GlcNac), mannose (Man), and glucose (Glc), having the stoichiometry GlcNac2Man9Glc3, is attached to the free amino group of a specific asparagine side chain. This carbohydrate assembly is called an N-linked glycan (Faye et al. 1992). The three terminal glucose residues are then removed by specific glucosidases, and the processed glycoprotein (i.e., a protein with covalently attached sugars) is ready for transport to the Golgi apparatus. The so-called N-linked glycoproteinsare then transported to the Golgi apparatus via small vesicles. The vesicles move through the cytosol and fuse with cisternae on the cis face of the Golgi apparatus (Figure 1.12).
Proteins and Polysaccharides for Secretion Are Processed in the Golgi Apparatus
The Golgi apparatus (also called Golgi complex) of plant cells is a dynamic structure consisting of one or more stacks of three to ten flattened membrane sacs, or cisternae, and an irregular network of tubules and vesicles called the trans Golgi network (TGN) (see Figure 1.12). Each individual stack is called a Golgi body or dictyosome. As Figure 1.12 shows, the Golgi body has distinct functional regions: The cisternae closest to the plasma membrane are called the trans face, and the cisternae closest to the center of the cell are called the cis face. The medial cisternae are between the trans and cis cisternae. The trans Golgi network is located on the trans face. The entire structure is stabilized by the presence of intercisternal elements, protein crosslinks that hold the cisternae together. Whereas in animal cells Golgi bodies tend to be clustered in one part of the cell and are interconnected via tubules, plant cells contain up to several hundred apparently separate Golgi bodies dispersed throughout the cytoplasm (Driouich et al. 1994). The Golgi apparatus plays a key role in the synthesis and secretion of complex polysaccharides (polymers composed of different types of sugars) and in the assembly of the oligosaccharide side chains of glycoproteins (Driouich et al. 1994). As noted already, the polypeptide chains of future glycoproteins are first synthesized on the rough ER, then transferred across the ER membrane, and glycosylated on the —NH2 groups of asparagine residues. Further modifications of, and additions to, the oligosaccharide side chains are carried out in the Golgi. Glycoproteins destined for secretion reach the Golgi via vesicles that bud off from the RER. The exact pathway of glycoproteins through the plant Golgi apparatus is not yet known. Since there appears to be no direct membrane continuity between successive cisternae, the contents of one cisterna are transferred to the next cisterna via small vesicles budding off from the margins, as occurs in the Golgi apparatus of animals. In some cases, however, entire cisternae may progress through the Golgi body and emerge from the trans face. Within the lumens of the Golgi cisternae, the glycoproteins are enzymatically modified. Certain sugars, such as mannose, are removed from the oligosaccharide chains, and other sugars are added. In addition to these modifications, glycosylation of the —OH groups of hydroxyproline, serine, threonine, and tyrosine residues (O-linked oligosaccharides) also occurs in the Golgi. After being processed within the Golgi, the glycoproteins leave the organelle in other vesicles, usually from the trans side of the stack. All of this processing appears to confer on each protein a specific tag or marker that specifies the ultimate destination of that protein inside or outside the cell. In plant cells, the Golgi body plays an important role in cell wall formation (see Chapter 15). Noncellulosic cell wall polysaccharides (hemicellulose and pectin) are synthesized, and a variety of glycoproteins, including hydroxyprolinerich glycoproteins, are processed within the Golgi. Secretory vesicles derived from the Golgi carry the polysaccharides and glycoproteins to the plasma membrane, where the vesicles fuse with the plasma membrane and empty their contents into the region of the cell wall. Secretory vesicles may either be smooth or have a protein coat. Vesicles budding from the ER are generally smooth. Most vesicles budding from the Golgi have protein coats of some type. These proteins aid in the budding process during vesicle formation. Vesicles involved in traffic from the ER to the Golgi, between Golgi compartments, and from the Golgi to the TGN have protein coats. Clathrin-coated vesicles (Figure 1.13) are involved in the transport of storage proteins from the Golgi to specialized protein-storing vacuoles. They also participate in endocytosis, the process that brings soluble and membrane-bound proteins into the cell. The
Central Vacuole Contains Water and Solutes
Mature living plant cells contain large, water-filled central vacuoles that can occupy 80 to 90% of the total volume of the cell (see Figure 1.4). Each vacuole is surrounded by a vacuolar membrane, or tonoplast. Many cells also have cytoplasmic strands that run through the vacuole, but each transvacuolar strand is surrounded by the tonoplast. In meristematic tissue, vacuoles are less prominent, though they are always present as small provacuoles. Provacuoles are produced by the trans Golgi network (see Figure 1.12). As the cell begins to mature, the provacuoles fuse to produce the large central vacuoles that are characteristic of most mature plant cells. In such cells, the cytoplasm is restricted to a thin layer surrounding the vacuole. The vacuole contains water and dissolved inorganic ions, organic acids, sugars, enzymes, and a variety of secondary metabolites (see Chapter 13), which often play roles in plant defense. Active solute accumulation provides the osmotic driving force for water uptake by the vacuole, which is required for plant cell enlargement. The turgor pressure generated by this water uptake provides the structural rigidity needed to keep herbaceous plants upright, since they lack the lignified support tissues of woody plants. Like animal lysosomes, plant vacuoles contain hydrolytic enzymes, including proteases, ribonucleases, and glycosidases. Unlike animal lysosomes, however, plant vacuoles do not participate in the turnover of macromolecules throughout the life of the cell. Instead, their degradative enzymes leak out into the cytosol as the cell undergoes senescence, thereby helping to recycle valuable nutrients to the living portion of the plant. Specialized protein-storing vacuoles, called protein bodies, are abundant in seeds. During germination the storage proteins in the protein bodies are hydrolyzed to amino acids and exported to the cytosol for use in protein synthesis. The hydrolytic enzymes are stored in specialized lytic vacuoles, which fuse with the protein bodies to initiate the breakdown process (Figure 1.14).
Mitochondria and Chloroplasts Are Sites of Energy Conversion
A typical plant cell has two types of energy-producing organelles: mitochondria and chloroplasts. Both types are separated from the cytosol by a double membrane (an outer and an inner membrane). Mitochondria (singular mitochondrion) are the cellular sites of respiration, a process in which the energy released from sugar metabolism is used for the synthesis of ATP (adenosine triphosphate) from ADP (adenosine diphosphate) and inorganic phosphate (Pi) (see Chapter 11). Mitochondria can vary in shape from spherical to tubular, but they all have a smooth outer membrane and a highly convoluted inner membrane (Figure 1.15). The infoldings of the inner membrane are called cristae (singular crista). The compartment enclosed by the inner membrane, the mitochondrial matrix, contains the enzymes of the pathway of intermediary metabolism called the Krebs cycle. In contrast to the mitochondrial outer membrane and all other membranes in the cell, the inner membrane of a mitochondrion is almost 70% protein and contains some phospholipids that are unique to the organelle (e.g., cardiolipin). The proteins in and on the inner membrane have special enzymatic and transport capacities. The inner membrane is highly impermeable to the passage of H+; that is, it serves as a barrier to the movement of protons. This important feature allows the formation of electrochemical gradients. Dissipation of such gradients by the controlled movement of H+ ions through the transmembrane enzyme ATP synthase is coupled to the phosphorylation of ADP to produce ATP. ATP can then be released to other cellular sites where energy is needed to drive specific reactions. Chloroplasts (Figure 1.16A) belong to another group of double membrane–enclosed organelles called plastids. Chloroplast membranes are rich in glycosylglycerides Chloroplast membranes contain chlorophyll and its associated proteins and are the sites of photosynthesis. In addition to their inner and outer envelope membranes, chloroplasts possess a third system of membranes called thylakoids. A stack of thylakoids forms a granum (plural grana) (Figure 1.16B). Proteins and pigments (chlorophylls and carotenoids) that function in the photochemical events of photosynthesis are embedded in the thylakoid membrane. The fluid compartment surrounding the thylakoids, called the stroma, is analogous to the matrix of the mitochondrion. Adjacent grana are connected by unstacked membranes called stroma lamellae (singular lamella). The different components of the photosynthetic apparatus are localized in different areas of the grana and the stroma lamellae. The ATP synthases of the chloroplast are located on the thylakoid membranes (Figure 1.16C). During photosynthesis, light-driven electron transfer reactions result in a proton gradient across the thylakoid membrane. As in the mitochondria, ATP is synthesized when the proton gradient is dissipated via the ATP synthase. Plastids that contain high concentrations of carotenoid pigments rather than chlorophyll are called chromoplasts. They are one of the causes of the yellow, orange, or red colors of many fruits and flowers, as well as of autumn leaves (Figure 1.17). Nonpigmented plastids are called leucoplasts. The most important type of leucoplast is the amyloplast, a starchstoring plastid. Amyloplasts are abundant in storage tissues of the shoot and root, and in seeds. Specialized amyloplasts in the root cap also serve as gravity sensors that direct root growth downward into the soil (see Chapter 19).
Mitochondria and Chloroplasts Are Semiautonomous Organelles
Both mitochondria and chloroplasts contain their own DNA and protein-synthesizing machinery (ribosomes, transfer RNAs, and other components) and are believed to have evolved from endosymbiotic bacteria. Both plastids and mitochondria divide by fission, and mitochondria can also undergo extensive fusion to form elongated structures or networks. The DNA of these organelles is in the form of circular chromosomes, similar to those of bacteria and very different from the linear chromosomes in the nucleus. These DNA circles are localized in specific regions of the mitochondrial matrix or plastid stroma called nucleoids. DNA replication in both mitochondria and chloroplasts is independent of DNAreplication in the nucleus. On the other hand, the numbers of these organelles within a given cell type remain approximately constant, suggesting that some aspects of organelle replication are under cellular regulation. The mitochondrial genome of plants consists of about 200 kilobase pairs (200,000 base pairs), a size considerably larger than that of most animal mitochondria. The mitochondria of meristematic cells are typically polyploid; that is, they contain multiple copies of the circular chromosome. However, the number of copies per mitochondrion gradually decreases as cells mature because the mitochondria continue to divide in the absence of DNA synthesis. Most of the proteins encoded by the mitochondrial genome are prokaryotic-type 70S ribosomal proteins and components of the electron transfer system. The majority of mitochondrial proteins, including Krebs cycle enzymes, are encoded by nuclear genes and are imported from the cytosol. The chloroplast genome is smaller than the mitochondrial genome, about 145 kilobase pairs (145,000 base pairs). Whereas mitochondria are polyploid only in the meristems, chloroplasts become polyploid during cell maturation. Thus the average amount of DNA per chloroplast in the plant is much greater than that of the mitochondria. The total amount of DNA from the mitochondria and plastids combined is about one-third of the nuclear genome (Gunning and Steer 1996). Chloroplast DNA encodes rRNA; transfer RNA (tRNA); the large subunit of the enzyme that fixes CO2, ribulose-1,5- bisphosphate carboxylase/oxygenase (rubisco); and several of the proteins that participate in photosynthesis. Nevertheless, the majority of chloroplast proteins, like those of mitochondria, are encoded by nuclear genes, synthesized in the cytosol, and transported to the organelle. Although mitochondria and chloroplasts have their own genomes and can divide independently of the cell, they are characterized as semiautonomous organelles because they depend on the nucleus for the majority of their proteins.
Different Plastid Types Are Interconvertible
Meristem cells contain proplastids, which have few or no internal membranes, no chlorophyll, and an incomplete complement of the enzymes necessary to carry out photosynthesis (Figure 1.18A). In angiosperms and some gymnosperms, chloroplast development from proplastids is triggered by light. Upon illumination, enzymes are formed inside the proplastid or imported from the cytosol, light-absorbing pigments are produced, and membranes proliferate rapidly, giving rise to stroma lamellae and grana stacks (Figure 1.18B). Seeds usually germinate in the soil away from light, and chloroplasts develop only when the young shoot is exposed to light. If seeds are germinated in the dark, the proplastids differentiate into etioplasts, which contain semicrystalline tubular arrays of membrane known as prolamellar bodies (Figure 1.18C). Instead of chlorophyll, the etioplast contains a pale yellow green precursor pigment, protochlorophyll. Within minutes after exposure to light, the etioplast differentiates, converting the prolamellar body into thylakoids and stroma lamellae, and the protochlorophyll into chlorophyll. The maintenance of chloroplast structure depends on the presence of light, and mature chloroplasts can revert to etioplasts during extended periods of darkness. Chloroplasts can be converted to chromoplasts, as in the case of autumn leaves and ripening fruit, and in some cases Another type of microbody, the glyoxysome, is present in oil-storing seeds. Glyoxysomes contain the glyoxylate cycle enzymes, which help convert stored fatty acids into sugars that can be translocated throughout the young plant to provide energy for growth (see Chapter 11). Because both types of microbodies carry out oxidative reactions, it has been suggested they may have evolved from primitive respiratory organelles that were superseded by mitochondria.
Oleosomes Are Lipid-Storing Organelles
In addition to starch and protein, many plants synthesize and store large quantities of triacylglycerol in the form of oil during seed development. These oils accumulate in organelles called oleosomes, also referred to as lipid bodies or spherosomes (Figure 1.20A). Oleosomes are unique among the organelles in that they are surrounded by a “half–unit membrane”—that is, a phospholipid monolayer—derived from the ER (Harwood 1997). The phospholipids in the half–unit membrane are oriented with their polar head groups toward the aqueous phase and their hydrophobic fatty acid tails facing the lumen, dissolved in the stored lipid. Oleosomes are thought to arise from the deposition of lipids within the bilayer itself (Figure 1.20B). Proteins called oleosins are present in the half–unit membrane (see Figure 1.20B). One of the functions of the oleosins may be to maintain each oleosome as a discrete organelle by preventing fusion. Oleosins may also help other proteins bind to the organelle surface. As noted earlier, during seed germination the lipids in the oleosomes are broken down and converted to sucrose with the help of the glyoxysome. The first step in the process is the hydrolysis of the fatty acid chains from the glycerol backbone by the enzyme lipase. Lipase is tightly associated with the surface of the half–unit membrane and may be attached to the oleosins.
THE CYTOSKELETON
The cytosol is organized into a three-dimensional network of filamentous proteins called the cytoskeleton. This network provides the spatial organization for the organelles and serves as a scaffolding for the movements of organelles and other cytoskeletal components. It also plays fundamental roles in mitosis, meiosis, cytokinesis, wall deposition, the maintenance of cell shape, and cell differentiation.
Plant Cells Contain Microtubules,Microfilaments, and Intermediate Filaments
Three types of cytoskeletal elements have been demonstrated in plant cells: microtubules, microfilaments, and intermediate filament–like structures. Each type is filamentous, having a fixed diameter and a variable length, up to many micrometers. Microtubules and microfilaments are macromolecular assemblies of globular proteins. Microtubules are hollow cylinders with an outer diameter of 25 nm; they are composed of polymers of the protein tubulin. The tubulin monomer of microtubules is a heterodimer composed of two similar polypeptide chains (á- and â-tubulin), each having an apparent molecular mass of 55,000 daltons (Figure 1.21A). A single microtubule consists of hundreds of thousands of tubulin monomers arranged in 13 columns called protofilaments. Microfilaments are solid, with a diameter of 7 nm; they are composed of a special form of the protein found in muscle: globular actin, or G-actin. Each actin molecule is composed of a single polypeptide with a molecular mass of approximately 42,000 daltons. Amicrofilament consists of two chains of polymerized actin subunits that intertwine in a helical fashion (Figure 1.21B). Intermediate filaments are a diverse group of helically wound fibrous elements, 10 nm in diameter. Intermediate filaments are composed of linear polypeptide monomers of various types. In animal cells, for example, the nuclear lamins are composed of a specific polypeptide monomer, while the keratins, a type of intermediate filament found in the cytoplasm, are composed of a different polypeptide monomer. In animal intermediate filaments, pairs of parallel monomers (i.e., aligned with their —NH2 groups at the same ends) are helically wound around each other in a coiled coil. Two coiled-coil dimers then align in an antiparallel fashion (i.e., with their —NH2 groups at opposite ends) to form a tetrameric unit. The tetrameric units then assemble into the final intermediate filament (Figure 1.22). Although nuclear lamins appear to be present in plant cells, there is as yet no convincing evidence for plant keratin intermediate filaments in the cytosol. As noted earlier, integral proteins cross-link the plasma membrane of plant cells to the rigid cell wall. Such connections to the wall undoubtedly stabilize the protoplast and help maintain cell shape. The plant cell wall thus serves as a kind of cellular exoskeleton, perhaps obviating the need for keratin-type intermediate filaments for structural support.
In the cell, actin and tubulin monomers exist as pools of free proteins that are in dynamic equilibrium with the polymerized forms. Polymerization requires energy: ATP is required for microfilament polymerization, GTP (guanosine triphosphate) for microtubule polymerization. The attachments between subunits in the polymer are noncovalent, but they are strong enough to render the structure stable under cellular conditions. Both microtubules and microfilaments are polarized; that is, the two ends are different. In microtubules, the polarity arises from the polarity of the á- and -tubulin heterodimer; in microfilaments, the polarity arises from the polarity of the actin monomer itself. The opposite ends of microtubules and microfilaments are termed plus and minus, and polymerization is more rapid at the positive end. Once formed, microtubules and microfilaments can disassemble. The overall rate of assembly and disassembly of these structures is affected by the relative concentrations of free or assembled subunits. In general, microtubules are more unstable than microfilaments. In animal cells, the half-life of an individual microtubule is about 10 minutes. Thus microtubules are said to exist in a state of dynamic instability. In contrast to microtubules and microfilaments, intermediate filaments lack polarity because of the antiparallel orientation of the dimers that make up the tetramers. In addition, intermediate filaments appear to be much more stable than either microtubules or microfilaments. Although very little is known about intermediate filament–like structures in plant cells, in animal cells nearly all of the intermediate- filament protein exists in the polymerized state. Microtubules Function in Mitosis and Cytokinesis Mitosis is the process by which previously replicated chromosomes are aligned, separated, and distributed in an orderly fashion to daughter cells (Figure 1.23). Microtubules are an integral part of mitosis. Before mitosis begins, microtubules in the cortical (outer) cytoplasm depolymerize, breaking down into their constituent subunits. The subunits then repolymerize before the start of prophase to form the preprophase band (PPB), a ring of microtubules encircling the nucleus (see Figure 1.23C–F). The PPB appears in the region where the future cell wall will form after the completion of mitosis, and it is thought to be involved in regulating the plane of cell division. During prophase, microtubules begin to assemble at two foci on opposite sides of the nucleus, forming the prophase spindle (Figure 1.24). Although not associated with any specific structure, these foci serve the same function as animal cell centrosomes in organizing and assembling microtubules. In early metaphase the nuclear envelope breaks down, the PPB disassembles, and new microtubules polymerize to form the mitotic spindle. In animal cells the spindle microtubules radiate toward each other from two discrete foci at the poles (the centrosomes), resulting in an ellipsoidal, or football-shaped, array of microtubules. The mitotic spindle of plant cells, which lack centrosomes, is more boxlike in shape because the spindle microtubules arise from a diffuse zone consisting of multiple foci at opposite ends of the cell and extend toward the middle in nearly parallel arrays (see Figure 1.24). Some of the microtubules of the spindle apparatus become attached to the chromosomes at their kinetochores, while others remain unattached. The kinetochores are located in the centromeric regions of the chromosomes. Some of the unattached microtubules overlap with microtubules from the opposite polar region in the spindle midzone. Cytokinesis is the process whereby a cell is partitioned into two progeny cells. Cytokinesis usually begins late in mitosis. The precursor of the new wall, the cell plate that Plant Cells 21 forms between incipient daughter cells, is rich in pectins (Figure 1.25). Cell plate formation in higher plants is a multistep process. Vesicle aggregation in the spindle midzone is organized by the phragmoplast, a complex of microtubules and ER that forms during late anaphase or early telophase from dissociated spindle subunits.
Microfilaments Are Involved in Cytoplasmic Streaming and in Tip Growth
Cytoplasmic streamingis the coordinated movement of particles and organelles through the cytosol in a helical path down one side of a cell and up the other side. Cytoplasmic streaming occurs in most plant cells and has been studied extensively in the giant cells of the green algae Chara and Nitella, in which speeds up to 75 ìm s–1 have been measured. The mechanism of cytoplasmic streaming involves bundles of microfilaments that are arranged parallel to the longitudinal direction of particle movement. The forces necessary for movement may be generated by an interaction of the microfilament protein actin with the protein myosin in a fashion comparable to that of the protein interaction that occurs during muscle contraction in animals. Myosins are proteins that have the ability to hydrolyze ATP to ADP and Pi when activated by binding to an actin microfilament. The energy released by ATP hydrolysis propels myosin molecules along the actin microfilament from the minus end to the plus end. Thus, myosins belong to the general class of motor proteins that drive cytoplasmic streaming and the movements of organelles within the cell. Examples of other motor proteins include the kinesins and dyneins, which drive movements of organelles and other cytoskeletal components along the surfaces of microtubules. Actin microfilaments also participate in the growth of the pollen tube. Upon germination, a pollen grain forms a tubular extension that grows down the style toward the embryo sac. As the tip of the pollen tube extends, new cell wall material is continually deposited to maintain the integrity of the wall. A network of microfilaments appears to guide vesicles containing wall precursors from their site of formation in the Golgi through the cytosol to the site of new wall formation at the tip. Fusion of these vesicles with the plasma membrane deposits wall precursors outside the cell, where they are assembled into wall material.
Intermediate Filaments Occur in the Cytosol and Nucleus of Plant Cells
Relatively little is known about plant intermediate filaments. Intermediate filament–like structures have been identified in the cytoplasm of plant cells (Yang et al. 1995), but these may not be based on keratin, as in animal cells, since as yet no plant keratin genes have been found. Nuclear lamins, intermediate filaments of another type that form a dense network on the inner surface of the nuclear membrane, have also been identified in plant cells (Frederick et al. 1992), and genes encoding laminlike proteins are present in the Arabidopsis genome. Presumably, plant lamins perform functions similar to those in animal cells as a structural component of the nuclear envelope.
CELL CYCLE REGULATION
The cell division cycle, or cell cycle, is the process by which cells reproduce themselves and their genetic material, the nuclear DNA. The four phases of the cell cycle are designated G1, S, G2, and M (Figure 1.26A).
Each Phase of the Cell Cycle Has a Specific Set of Biochemical and Cellular Activities
Nuclear DNA is prepared for replication in G1 by the assembly of a prereplication complex at the origins of replication along the chromatin. DNA is replicated during the S phase, and G2 cells prepare for mitosis. The whole architecture of the cell is altered as cells enter mitosis: The nuclear envelope breaks down, chromatin condenses to form recognizable chromosomes, the mitotic spindle forms, and the replicated chromosomes attach to the spindle fibers. The transition from metaphase to anaphase of mitosis marks a major transition point when the two chromatids of each replicated chromosome, which were held together at their kinetochores, are separated and the daughter chromosomes are pulled to opposite poles by spindle fibers. At a key regulatory point early in G1 of the cell cycle, the cell becomes committed to the initiation of DNA synthesis. In yeasts, this point is called START. Once a cell has passed START, it is irreversibly committed to initiating DNA synthesis and completing the cell cycle through mitosis and cytokinesis. After the cell has completed mitosis, it may initiate another complete cycle (G1 through mitosis), or it may leave the cell cycle and differentiate. This choice is made at the critical G1 point, before the cell begins to replicate its DNA. DNAreplication and mitosis are linked in mammalian cells. Often mammalian cells that have stopped dividing can be stimulated to reenter the cell cycle by a variety of hormones and growth factors. When they do so, they reenter the cell cycle at the critical point in early G1. In contrast, plant cells can leave the cell division cycle either before or after replicating their DNA (i.e., during G1 or G2). As a consequence, whereas most animal cells are diploid (having two sets of chromosomes), plant cells frequently are tetraploid (having four sets of chromosomes), or even polyploid (having many sets of chromosomes), after going through additional cycles of nuclear DNA replication without mitosis.
The Cell Cycle Is Regulated by Protein Kinases
The mechanism regulating the progression of cells through their division cycle is highly conserved in evolution, and plants have retained the basic components of this mechanism (Renaudin et al. 1996). The key enzymes that control the transitions between the different states of the cell cycle, and the entry of nondividing cells into the cell cycle, are the cyclin-dependent protein kinases, or CDKs (Figure 1.26B). Protein kinases are enzymes that phosphorylate proteins using ATP. Most multicellular eukaryotes use several protein kinases that are active in different phases of the cell cycle. All depend on regulatory subunits called cyclins for their activities. The regulated activity of CDKs is essential for the transitions from G1 to S and from G2 to M, and for the entry of nondividing cells into the cell cycle. CDK activity can be regulated in various ways, but two of the most important mechanisms are (1) cyclin synthesis and destruction and (2) the phosphorylation and dephosphorylation of key amino acid residues within the CDK protein. CDKs are inactive unless they are associated with a cyclin. Most cyclins turn over rapidly. They are synthesized and then actively degraded (using ATP) at specific points in the cell cycle. Cyclins are degraded in the cytosol by a large proteolytic complex called the proteasome. Before being degraded by the proteasome, the cyclins are marked for destruction by the attachment of a small protein called ubiquitin, a process that requires ATP. Ubiquitination is a general mechanism for tagging cellular proteins destined for turnover (see Chapter 14). The transition from G1 to S requires a set of cyclins (known as G1 cyclins) different from those required in the transition from G2 to mitosis, where mitotic cyclins activate the CDKs (see Figure 1.26B). CDKs possess two tyrosine phosphorylation sites: One causes activation of the enzyme; the other causes inactivation. Specific kinases carry out both the stimulatory and the inhibitory phosphorylations. Similarly, protein phosphatases can remove phosphate from CDKs, either stimulating or inhibiting their activity, depending on the position of the phosphate. The addition or removal of phosphate groups from CDKs is highly regulated and an important mechanism for the control of cell cycle progression (see Figure 1.26B). Cyclin inhibitors play an important role in regulating the cell cycle in animals, and probably in plants as well, although little is known about plant cyclin inhibitors. Finally, as we will see later in the book, certain plant hormones are able to regulate the cell cycle by regulating the synthesis of key enzymes in the regulatory pathway.
PLASMODESMATA
Plasmodesmata(singular plasmodesma) are tubular extensions of the plasma membrane, 40 to 50 nm in diameter, that traverse the cell wall and connect the cytoplasms of adjacent cells. Because most plant cells are interconnected in this way, their cytoplasms form a continuum referred to as the symplast. Intercellular transport of solutes through plasmodesmata is thus called symplastic transport (see Chapters 4 and 6).
There Are Two Types of Plasmodesmata: Primary and Secondary
Primary plasmodesmataform during cytokinesis when Golgi-derived vesicles containing cell wall precursors fuse to form the cell plate (the future middle lamella). Rather than forming a continuous uninterrupted sheet, the newly deposited cell plate is penetrated by numerous pores (Figure 1.27A), where remnants of the spindle apparatus, consisting of ER and microtubules, disrupt vesicle fusion. Further deposition of wall polymers increases the thickness of the two primary cell walls on either side of the middle lamella, generating linear membrane-lined channels (Figure 1.27B). Development of primary plasmodesmata thus provides direct continuity and communication between cells that are clonally related (i.e., derived from the same mother cell). Secondary plasmodesmata form between cells after their cell walls have been deposited. They arise either by evagination of the plasma membrane at the cell surface, or by branching from a primary plasmodesma (Lucas and Wolf 1993). In addition to increasing the communication between cells that are clonally related, secondary plasmodesmata allow symplastic continuity between cells that are not clonally related.
Plasmodesmata Have a Complex Internal Structure
Like nuclear pores, plasmodesmata have a complex internal structure that functions in regulating macromolecular traffic from cell to cell. Each plasmodesma contains a narrow tubule of ER called a desmotubule (see Figure 1.27). The desmotubule is continuous with the ER of the adjacent cells. Thus the symplast joins not only the cytosol of neighboring cells, but the contents of the ER lumens as well. However, it is not clear that the desmotubule actually represents a passage, since there does not appear to be a space between the membranes, which are tightly appressed. Globular proteins are associated with both the desmotubule membrane and the plasma membrane within the pore (see Figure 1.27B). These globular proteins appear to be interconnected by spokelike extensions, dividing the pore into eight to ten microchannels (Ding et al. 1992). Some molecules can pass from cell to cell through plasmodesmata, probably by flowing through the microchannels, although the exact pathway of communication has not been established. By following the movement of fluorescent dye molecules of different sizes through plasmodesmata connecting leaf epidermal cells, Robards and Lucas (1990) determined the limiting molecular mass for transport to be about 700 to 1000 daltons, equivalent to a molecular size of about 1.5 to 2.0 nm. This is the size exclusion limit, or SEL, of plasmodesmata. If the width of the cytoplasmic sleeve is approximately 5 to 6 nm, how are molecules larger than 2.0 nm excluded? The proteins attached to the plasma membrane and the ER within the plasmodesmata appear to act to restrict the size of molecules that can pass through the pore. As we’ll see in Chapter 16, the SELs of plasmodesmata can be regulated. The mechanism for regulating the SEL is poorly understood, but the localization of both actin and myosin within plasmodesmata, possibly forming the “spoke” extensions (see Figure 1.27B), suggests that they may participate in the process (White et al. 1994; Radford and White 1996). Recent studies have also implicated calcium-dependent protein kinases in the regulation of plasmodesmatal SEL.
SUMMARY
Despite their great diversity in form and size, all plants carry out similar physiological processes. As primary producers, plants convert solar energy to chemical energy. Being nonmotile, plants must grow toward light, and they must have efficient vascular systems for movement of water, mineral nutrients, and photosynthetic products throughout the plant body. Green land plants must also have mechanisms for avoiding desiccation. The major vegetative organ systems of seed plants are the shoot and the root. The shoot consists of two types of organs: stems and leaves. Unlike animal development, plant growth is indeterminate because of the presence of permanent meristem tissue at the shoot and root apices, which gives rise to new tissues and organs during the entire vegetative phase of the life cycle. Lateral meristems (the vascular cambium and the cork cambium) produce growth in girth, or secondary growth. Three major tissue systems are recognized: dermal, ground, and vascular. Each of these tissues contains a variety of cell types specialized for different functions. Plants are eukaryotes and have the typical eukaryotic cell organization, consisting of nucleus and cytoplasm. The nuclear genome directs the growth and development of the organism. The cytoplasm is enclosed by a plasma membrane and contains numerous membrane-enclosed organelles, including plastids, mitochondria, microbodies, oleosomes, and a large central vacuole. Chloroplasts and mitochondria are semiautonomous organelles that contain their own DNA. Nevertheless, most of their proteins are encoded by nuclear DNA and are imported from the cytosol. The cytoskeletal components—microtubules, microfilaments, and intermediate filaments—participate in a variety of processes involving intracellular movements, such as mitosis, cytoplasmic streaming, secretory vesicle transport transport, cell plate formation, and cellulose microfibril deposition. The process by which cells reproduce is called the cell cycle. The cell cycle consists of the G1, S, G2, and M phases. The transition from one phase to another is regulated by cyclin-dependent protein kinases. The activity of the CDKs is regulated by cyclins and by protein phosphorylation. During cytokinesis, the phragmoplast gives rise to the cell plate in a multistep process that involves vesicle fusion. After cytokinesis, primary cell walls are deposited. The cytosol of adjacent cells is continuous through the cell walls because of the presence of membrane-lined channels called plasmodesmata, which play a role in cell–cell communication-
Plant cells typically have a cell wall surrounding the protoplasm, which consists of all the living components of a cell. These living components are bounded by a membrane called the plasma membrane. All cellular components between the plasma membrane and a relatively large body called the nucleusare known as cytoplasm. Within the cytoplasm is a souplike fluid called cytosol, in which various bodies called organelles are dispersed. Organelles are persistent structures of various shapes and sizes with specialized functions in the cell; most, but not all, are bounded by membranes
playlist- Biochemistry – Carbohydrates, Lipids and Cell Membrane
The main structural component of cell walls is cellulose, which is composed of 100 to 15,000 glucose monomers in long chains, and is the most abundant polymer on earth. As a primary food source for grazing animals and at least indirectly for nearly all other living organisms, it could be said that most life on earth relies directly or indirectly on the cell wall. Humans also depend on cell walls because they provide clothing, shelter, furniture, paper, and fuel.
In addition to cellulose, cell walls typically contain a matrix of hemicellulose (a gluelike substance that holds cellulose fibrils together), pectin (the organic material that gives stiffness to fruit jellies), and glycoproteins (proteins that have sugars associated with their molecules). A middle lamella, which consists of a layer of pectin, is first produced when new cell walls are formed. This middle lamella is normally shared by two adjacent cells and is so thin that it may not be visible with an ordinary light microscope unless it is specially stained. A flexible primary wall, consisting of a fine network of cellulose, hemicellulose, pectin, and glycoproteins, is laid down on either side of the middle lamella Reorganization, synthesis of new molecules, and insertion of new wall polymers lead to – rearrangement of the cell wall during growth. Secondary walls, which are produced inside the primary walls, are derived from primary walls by thickening and inclusion of lignin, a complex polymer. Secondary cell walls of plants generally contain more cellulose (40% to 80%) than primary walls. As the cell ages, wall thickness can vary, occupying as little as 5% to more than 95% of the volume of the cells. During secondary wall formation, cellulose microfibrils become embedded in lignin, much like steel rods are embedded in concrete to form prestressed concrete
The Plasma Membrane
The outer boundary of the living part of the cell, the plasma membrane, [is] composed of phospholipids arranged in two layers, with proteins interspersed throughout This fluid mosaic model for the plasma membrane implies a dynamic structure with numerous components, some of which can migrate and interact directly with each other. Covalent bonds link carbohydrates to both lipids and proteins on the outer surfaces of membranes
The nucleus is the control center of the cell Each nucleus is bounded by two membranes, which together constitute the nuclear envelope. The nucleus contains a granular-appearing fluid called nucleoplasm, which is packed with short fibers that are about 10 nanometers in diameter; several different larger bodies are suspended within it. Of the larger nuclear bodies, the most noticeable are one or more nucleoli (singular: nucleolus), which are composed primarily of RNA and associated proteins. The nucleus includes thin strands of chromatin. When a nucleus divides, the chromatin strands coil, becoming shorter and thicker, and in their condensed condition, they are called chromosomes. Chromatin is composed of protein and DNA
The Endoplasmic Reticulum
The outer membrane of the nucleus is connected and continuous with the endoplasmic reticulum. The endoplasmic reticulum facilitates cellular communication and channeling of materials.
Ribosomal subunits are assembled within the nucleolus, released, and in association with special RNA molecules, they initiate protein synthesis. Once assembled, complete ribosomes may line the outside of the endoplasmic reticulum but can also occur unattached in the cytoplasm, chloroplasts, or other organelles.
Stacks of flattened discs or vesicles known as dictyosomes may be scattered throughout the cytoplasm of a cell. Dictyosomes are often bounded by branching tubules that originate from the endoplasmic reticulum, but are not directly connected to it Dictyosomes are involved in the modification of carbohydrates attached to proteins that are synthesized and packaged in the endoplasmic reticulum. Complex polysaccharides are also assembled within the dictyosomes and collect in small vesicles (tiny, blisterlike bodies) that are pinched off from the margins. These vesicles migrate to the plasma membrane, fuse with it, and secrete their contents outside of the cell. Substances secreted by vesicles may include cell-wall polysaccharides, floral nectars, and essential oils found in herbs.
Seventy-five to 125 is quite common, with the green cells of a few plants having up to several hundred.
Within the chloroplast are numerous grana (singular: granum), which are formed from membranes and have the appearance of stacks of coins with double membranes. There are usually about 40 to 60 grana linked together by arms in each chloroplast, and each granum may contain from two or three to more than 100 stacked thylakoids. In reality, thylakoids are part of an overlapping and continuous membrane system suspended in the liquid portion of the chloroplast
The thylakoid membranes contain greenchlorophyll and other pigments. These “coin stacks” of grana are vital to life as we know it, for it is within the thylakoids that the first steps of the important process of photosynthesis The liquid portion of the chloroplast is a colorless fluid matrix called stroma, which contains enzymes involved in photosynthesis.
Chromoplasts are another type of plastid found in some cells of more complex plants. Although chromoplasts are similar to chloroplasts in size, they vary considerably in shape, often being somewhat angular. They sometimes develop from chloroplasts through internal changes that include the disappearance of chlorophyll. Chromoplasts are yellow, orange, or red in color due to the presence of carotenoid pigments, which they synthesize and accumulate. They are most abundant in the yellow, orange, or some red parts of plants, such as ripe tomatoes, carrots, or red peppers These carotenoid pigments, which are lipid soluble, are not, however, the predominant pigments in most red flower petals. The anthocyanin pigments of most red flower petals are water soluble.
Leucoplasts are yet another type of plastid common to cells of higher plants. They are essentially colorless and include amyloplasts, which synthesize starches, and elaioplasts, which synthesize oils. If exposed to light, some leucoplasts will develop into chloroplasts, and vice versa.
Plastids of all types develop from proplastids
.
Mitochondria
Mitochondria (singular: mitochondrion) are often referred to as the powerhouses of the cell, for it is within them that energy is released from organic molecules by the process of cellular respiration
Carbon skeletons and fatty acid chains are also rearranged within mitochondria, allowing for the building of a wide variety of organic molecules.
Microbodies
Various small bodies distributed throughout the cytoplasm tend to give it a granular appearance. Examples of such components include types of small, spherical organelles called microbodies, which contain specialized enzymes and are bounded by a single membrane. Peroxisomes, for instance, contain enzymes needed by some plants to survive during hot conditions in a process called photorespiration
Vacuoles
In a mature living plant cell, as much as 90% or more of the volume may be taken up by one or two large central vacuoles that are bounded by vacuolar membranes (tonoplasts) The vacuolar membranes, which constitute the inner boundaries of the living part of the cell, are similar in structure and function
to plasma membranes.
The Cytoskeleton
The cytoskeleton is involved in movement within a cell and in a cell’s architecture. It is an intricate network con-structed mainly of two kinds of fibers—microtubules and –microfilaments.
Microtubules control the addition of cell-ulose to the cell wall . They are also involved in cell division, movement of cytoplasmic organelles, controlling the movement of vesicles containing cell-wall components assembled by dictyosomes, and movement of the tiny whiplike flagella and cilia possessed by some cells Microtubules are unbranched, thin, hollow, tubelike structures that resemble tiny straws. They are composed of proteins called tubulins and are of varying lengths, most being between 15 and 25 nanometers in diameter. They are most commonly found just inside the plasma membrane. Microtubules are also found in the special fibers that form the spindles and phragmoplasts of dividing cells
Microfilaments, which play a major role in the contraction and movement of cells in multicellular animals, are present in nearly all cells. They are three or four times thinner than microtubules and consist of long, fine threads of protein with an average diameter of 6 nanometers. They are often in bundles and appear to play a role in the cytoplasmic streaming (sometimes referred to as cyclosis) that occurs in all living cells. When cytoplasmic streaming is occurring, a microscope reveals the apparent movement of organelles as a current within the cytoplasm carries them around within the walls. This streaming probably facilitates exchanges of materials within the cell and plays a role in the movement of substances from cell to cell. The precise nature and origin of cytoplasmic streaming is still not known, but there is evidence that bundles of microfilaments may be responsible for it. Other evidence suggests that it may be related to the transport of cellular substances by microtubules.
CELLULAR REPRODUCTION
The Cell Cycle
When cells divide, they go through an orderly series of events known as the cell cycle. This cycle is usually divided into interphase and mitosis, mitosis itself being subdivided into four phases
videos- Cell Cycle and Interphase, Cytokinesis: Plant vs. Animal Cells, phases of mitosis
Interphase
Living cells that are not dividing are said to be in interphase, a period during which chromosomes are not visible with light microscopes. It is such cells that have been discussed up to this point. For many years, immature cells were considered to be “resting” when they were not actually dividing, but we know now that three consecutive periods of intense activity take place during interphase. These intervals are designated as gap (or growth) 1, synthesis, and gap (or growth) 2 periods, usually referred to as G1, S, and G2, respectively.
Also, ribosomes, RNA, and substances that either inhibit or stimulate the S period that follows are produced. During the S period, the unique process of DNA replication (duplication) takes place. In the G2 period, mitochondria and other organelles divide, and microtubules and other substances directly involved in mitosis are produced. Coiling and condensation of chromosomes also begin during G2.
Mitosis
Strictly speaking, mitosis refers to the division of the nucleus alone, but with a few exceptions seen in algae and fungi the division of the remainder of the cell, called cytokinesis, normally accompanies or follows mitosis. Both processes will be considered together here.
In flowering plants, conifers, and other higher plants, mitosis occurs in specific regions, or tissues, called meri-stems ( Meristems are found in the root and stem tips and also in a thin, perforated, and branching cylinder of tissue called the vascular cambium (often referred to simply as the cambium), located in the interior of some stems and roots a short distance from the surface. In some herbaceous and most woody plants, a second meristem similar in form to the cambium lies between the cambium and the outer bark. This second meristem is called the cork cambium
Mitosis is initiated with the appearance of a ringlike preprophase band of microtubules just beneath the plasma membrane and isusually divided into four arbitrary phases
Prophase
The main features of prophase are (1) the chromosomes become shorter and thicker, and their two-stranded nature becomes apparent; (2) the nuclear envelope fragments, and the nucleolus disintegrates.
.Metaphase
The main feature of metaphase is the alignment of the chromosomes in a circle midway between the two poles around the circumference of the spindle and in the same plane as that previously occupied by the preprophase band. This invisible circular plate is perpendicular to the axis of the spindle and is something like the equator of a globe.
Anaphase
Anaphase—the briefest of the phases—involves the sister chromatids of each chromosome separating and moving to opposite poles
Telophase
The five main features of telophase are (1) each group of daughter chromosomes becomes surrounded by a reformed nuclear envelope; (2) the daughter chromosomes become longer and thinner and finally become indistinguishable; (3) nucleoli reappear; (4) many of the spindle fibers disintegrate; and (5) a cell plate forms.
HIGHER PLANT CELLS VERSUS ANIMAL CELLS
All animals have either internal or external skeletons or skeleton-like systems to support their tissues. Animal cells do not have cell walls; instead, the plasma membrane, called the cell membrane by most zoologists (animal scientists), forms the outer boundary of animal cells. Higher plant cells have walls that are thickened and rigid to varying degrees, with a framework of cellulose fibrils. Higher plant cells also have plasmodesmata connecting the protoplasts with each other through microscopic holes in the walls. Animal cells lack plasmodesmata since they have no walls. When higher plant cells divide, a cell plate is formed during the telophase of mitosis, but cell plates do not form in animal cells, which divide by pinching in two.
Summary
All living organisms are composed of cells. Cells are modified according to the functions they perform; some live for a few days, while others live for many years.
The discovery of cells is associated with the development of the microscope. In 1665, Robert Hooke coined the word cells for boxlike compartments he saw in cork. Leeuwenhoek and Grew reported frequently during the next 50 years on the existence ofcells in a variety of tissues.
In 1809, Lamarck concluded that all living tissue is composed of cells, and in 1824, Dutrochet reinforced Lamarck’s conclusions. In 1833, Brown discovered that all cells contain a nucleus, and shortly thereafter, Schleiden saw a nucleolus within a nucleus. Schleiden and Schwann are credited with developing the cell theory in 1838 to 1839. The theory holds that all living organisms are composed of cells and that cells form a unifying, structural basis of organization.
In 1858, Virchow contended that every cell comes from a preexisting cell and that there is no spontaneous generation of cells from dust. In 1862, Pasteur experimentally confirmed Virchow’s contentions and later proved that fermentation involves activity of yeast cells. In 1897, Buchner found that yeast cells do not need to be alive for fermentation to occur. This led to the discovery of enzymes.
Light microscopes can magnify up to 1,500 times. Thinly sliced materials can be viewed with compound microscopes. Opaque objects can be viewed with stereo-microscopes; most magnify up to 30 times.
Electron microscopes have electromagnetic lenses and a beam of electrons within a vacuum that achieve magnification. Transmission electron microscopes magnify up to 200,000 or more times. Scanning electron microscopes, which can be used with opaque objects, usually magnify up to 10,000 times. 7. Scanning tunneling microscopes use a minute probe to scan surfaces at a width as narrow as that of two atoms.
Eukaryotic cells are the subject of this topc. Pro-karyotic cells, lack some of the features of eukaryotic cell.
Cells are minute, varying in diameter between 10 and 100 micrometers. They number into the billions in larger organisms, such as trees. Plant cells are bounded by walls that surround the living protoplast. The cytoplasm contains a souplike fluid called the cytoso and all cellular components between the plasma membrane and nucleus.
A pectic middle lamella is sandwiched between the primary cell walls of adjacent cells. The primary wall and also the secondary cell wall, often added inside the primary wall, are composed of cellulose polymers, with hemicelluloses and glycoproteins. Secondary cell walls contain lignin, which strengthens the wall.
Living cells are in contact with one another via fine strands of cytoplasm called plasmodesmata, which often extend through minute holes in the walls.
A flexible plasma membrane, which is sandwich-like and often forms folds, constitutes the outer boundary of the cytoplasm. It regulates the substances that enter and leave the cell.
The nucleus is bounded by a nuclear envelope consisting of two membranes that are perforated by numerous pores. Within the nucleus are a fluid called nucleoplasm, one or more spherical nucleoli, and thin strands of chromatin, which condense and become chromosomes when nuclei divide. Each species of organism has a specific number of chromosomes in each cell.
The endoplasmic reticulum is a system of flattened sacs and tubes associated with the storing and transporting of protein and other cell products. Granular particles called ribosomes, which function in protein synthesis, may line the outer surfaces of the endoplasmic reticulum. Ribosomes also occur independently in the cytoplasm.
Dictyosomes are structures that appear as stacks of sacs and function as collecting and packaging centers for the cell.
Plastids are larger green, orange, red, or colorless organelles. Green plastids, known as chloroplasts, contain enzymes that catalyze reactions of photosynthesis. These reactions take place in the membranes of structures that resemble stacks of coins, called thylakoids, as well as the surrounding matrix, called the stroma. Plastids develop from proplastids, which divide frequently, and also arise from the division of mature plastids.
Mitochondria are tiny, numerous organelles that are bounded by two membranes with inner platelike folds called cristae; they are associatedwith cellular respiration.
One or more vacuoles may occupy 90% or more of the volume of a mature cell. Vacuoles are bounded by a vacuolar membrane (tonoplast) and contain a watery fluid called cell sap. Cell sap contains dissolved substances and sometimes water-soluble red or blue anthocyanin pigments.
The cytoskeleton, which is involved in the architecture of cells and internal movement, is composed of microtubules and microfilaments. Microfilaments may be responsible for cytoplasmic streaming.
Cells that are not dividing are in interphase, which is subdivided into three periods of intense activity that precede mitosis or division of the nucleus. Mitosis is usually accompanied by division of the rest of the cell and takes place in meristems.
Mitosis is arbitrarily divided into four phases: (1) prophase, in which the chromosomes and their two-stranded nature become apparent and the nuclear envelope breaks down; (2) metaphase, in which the chromosomes become aligned at the equator of the cell; a spindle composed of spindle fibers is fully developed, with some spindle fibers being attached to the chromosomes at their centromeres; (3) anaphase, in which the sister chromatids of each chromosome (now called daughter chromosomes) separate lengthwise, with each group of daughter chromosomes migrating to opposite poles of the cell; and (4) telophase, in which each group of daughter chromosomes becomes surrounded by a nuclear envelope, thus becoming new nuclei, and a wall dividing the daughter nuclei forms, creating two daughter cells.
Animal cells differ from those of higher plants in not having a wall, plastids, or large vacuoles. Also, they have keg-shaped centrioles in pairs just outside the nucleus and pinch in two instead of forming a cell plate when they divide.
Tissues
MERISTEMATIC TISSUES
Unlike animals, plants have permanent regions of growth called meristems, or meristematic tissues, where cells actively divide
Apical Meristems
Apical meristems are meristematic tissues found at, or near, the tips of roots and shoots, which increase in length as the apical meristems produce new cells. This type of growth is known as primary growth. Three primary meristems, as well as embryo leaves and buds, develop from apical meristems. These primary meristems are called protoderm, ground meristem, and procambium. The tissues they produce are called primary tissues.
Lateral Meristems
The vascular cambium and cork cambium, discussed next, are lateral meristems, which produce tissues that increase the girth of roots and stems. Such growth is termed secondary growth.
Vascular Cambium
The vascular cambium, often referred to simply as the cambium, produces secondary tissues that function primarily in support and conduction. The cambium, which extends throughout the length of roots and stems in perennial and many annual plants, is in the form of a thin cylinder of mostly brick-shaped cells. The cambial cylinder often branches, except at the tips, and the tissues it produces are responsible for most of the increase in a plant’s girth as it grows. The individual remaining cells of the cambium are referred to as initials, while their sister cells are called derivatives.
Cork Cambium
The cork cambium, like the vascular cambium, is in the form of a thin cylinder that runs the length of roots and stems of woody plants. It lies outside of the vascular cambium, just inside the outer bark, which it produces. The tissues laid down by the vascular cambium and the cork cambium are called secondary tissues, since they are produced after the primary tissues have matured
Intercalary Meristems
Grasses and related plants have neither a vascular cambium nor a cork cambium. They do, however, have apical meri-stems, and, in the vicinity of nodes (leaf attachment areas), they have other meristematic tissues called intercalary meristems. The intercalary meristems develop at intervals along stems, where, like the tissues produced by apical meristems, their tissues add to stem length.
Simple Tissues
Parenchyma
Parenchyma tissue is composed of parenchyma cells ( They are more or less spherical in shape when they are first produced, but when all the parenchyma cells push up against one another, their thin, pliable walls are flattened at the points of contact. As a result, parenchyma cells assume various shapes and sizes, with the majority having 14 sides. They tend to have large vacuoles and may contain starch grains, oils, tannins (tanning or dyeing substances), crystals, and various other secretions. More often than not, parenchyma cells have spaces between them; in fact, in water lilies and other aquatic plants, the intercellular spaces are quite extensive and form a network throughout the entire plant. This type of parenchyma tissue—with extensive connected air spaces—is referred to as aerenchyma.
Parenchyma cells containing numerous chloroplasts (as found in leaves) are collectively referred to as chlorenchyma tissue. Chlorenchyma tissues function mainly in photosynthesis, while parenchyma tissues without chloroplasts function mostly in food or water storage. For example, the soft, edible parts of most fruits and vegetables consist largely of parenchyma.
Collenchyma
Collenchyma cells like parenchyma cells, have living cytoplasm and may remain alive a long time. Their walls generally are thicker and more uneven in thickness than those of parenchyma cells. The unevenness is due to extra primary wall in the corners. Collenchyma cells often occur just beneath the epidermis; typically, they are longer than they are wide, and their walls are pliable as well as strong. They provide flexible support for both growing organs and mature organs, such
as leaves and floral parts. The “strings” of celery that get stuck in our teeth, for example, are composed of collenchyma cells
Sclerenchyma
Sclerenchyma tissue consists of cells that have thick, tough, secondary walls, normally impregnated with lignin. Most sclerenchyma cells are dead at maturity and function in support. Two forms of sclerenchyma occur: sclereids and fibers. Sclereids may be randomly distributed in other tissues. For example, the slightly gritty texture of pears is due to the presence of groups of sclereids, or stone cells, as they are sometimes called.
Xylem tissue is an important component of the “plumbing” and storage systems of a plant and is the chief conducting tissue throughout all organs for water and minerals absorbed by the roots. Xylem consists of a combination of parenchyma cells, fibers, vessels, tracheids, and ray cells (Vessels are long tubes composed of individual cells called vessel elements that are open at each end. As each vessel element develops, the perforation plate, in some instances, can become barlike strips of wall material that extend across the openings. However, the flow of fluid through the vessels is not blocked by the strips.
Tracheids, which, like vessel elements, are dead at maturity and have relatively thick secondary cell walls, are tapered at each end, the ends overlapping with those of other tracheids. Tracheids have no openings similar to those of vessels, but there are usually pairs of pits present wherever two tracheids are in contact with one another. Pits are areas in which no secondary wall material has been deposited and, they allow water to pass from cell to cell. Figure 4.8 illustrates how, in some plants, pit pairs function in regulating the passage of water between adjacent cells. In cone-bearing trees and certain other non-flowering plants, the xylem is composed almost entirely of tracheids. The walls of many tracheids, as well as vessel elements, have spiral thickenings on them that are easily seen with the light microscope Most conduction through xylem is upward, but some is lateral (sideways). The lateral conduction takes place in the rays. Ray cells, which also function in food storage, are actually long-lived parenchyma cells that are produced in horizontal rows by special ray initials of the vascular cambium. In woody plants, the rays radiate out from the center of stems and roots like the spokes of a wheel.
Phloem
Phloem tissue which conducts dissolved food materials (primarily sugars) produced by photosynthesis throughout the plant, is composed mostly of two types of cells without secondary walls. The relatively large, more or less cylindrical sieve tube members have narrower, more tapered companion cells closely associated with them. Phloem is derived from the parent cells of the cambium, which also produce xylem cells; it often also includes fibers, parenchyma, and ray cells. Sieve tube members, like vessel elements, are laid end to end, forming sieve tubes. Unlike vessel elements, however, the end walls have no large openings; instead, the walls are full of small pores through which the cytoplasm extends from cell to cell. These porous regions of sieve tube members are called sieve plates. Sieve tube members have no nuclei at maturity, even though their cytoplasm is very active in the conduction of food materials in solution throughout the plant. Apparently, the adjacent companion cells form a very close relationship with the sieve tubes next to them and aid in the conduction of the food. Living sieve tube members contain a polymer called callose that stays in solution as long as the cell contents are under pressure. If an insect such as an aphid injures a cell, however, the pressure drops, and the callose precipitates. The callose and a phloem protein are then carried to the nearest sieve plate where they form a callus plug that prevents leaking of the sieve tube contents.
Sieve cells, which are found in ferns and cone-bearing trees, are similar to sieve tube members but tend to overlap at their ends rather than form continuous tubes. Like sieve tube members, they have no nuclei at maturity, but they have no adjacent companion cells. They do have adjacent albuminous cells, which are equivalent to companion cells and apparently function in the same manner.
The outermost layer of cells of all young plant organs is called the epidermis. Since it is in direct contact with the environment, it is subject to modification by the environment and often includes several different kinds of cells. The epidermis is usually one cell thick, but a few plants produce aerial roots called velamen roots (e.g., orchids) in which the epidermis may be several cells thick, with the outer cells functioning something like a sponge. Such a multiple-layered epidermis also occurs in the leaves of some tropical figs and members of the Pepper Family (Piperaceae), where it protects a plant from desiccation. Most epidermal cells secrete a fatty substance called cutin within and on the surface of the outer walls. Cutin forms a protective layer called the cuticle The thickness of the cuticle (or, more importantly, wax secreted on top of the cuticle by the epidermis) to a large extent determines how much water is lost through the cell walls by evaporation. The cuticle is also exceptionally resistant to bacteria and other disease organisms and has been recovered from fossil plants millions of years old. The waxes deposited on the cuticle in a number of plants can reach the surface by diffusion, migrate between cells, or travel through microscopic channels in the cell walls. The susceptibility of a plant to herbicides may depend on the thickness of these wax layers. Some wax deposits are extensive enough to have commercial value. Carnauba wax, for example, is deposited on the leaves of the wax palm. It and other waxes are harvested for use in polishes and, in the past, for phonograph records. In colonial times, a wax obtained from boiling leaves and fruits of the wax myrtle was used to make bayberry candles. In leaves, the epidermal cell walls perpendicular to the surface often assume bizarre shapes that, under the microscope, give them the appearance of pieces of a jigsaw puzzle. Epidermal cells of roots produce tubular extensions called root hairs a short distance behind the growing tips. The root hairs greatly increase the absorptive area of the surface. Hairs of a different nature occur on the epidermis of above-ground parts of plants. These hairs form outgrowths consisting of one to several cells Leaves also have numerous small pores, the stomata, bordered by pairs of specialized epidermal cells called guard cells Guard cells differ in shape from other epidermal cells; they also differ in that chloroplasts are present within themSome epidermal cells may be modified as glands that secrete protective or other substances, or modified as hairs that either reduce water loss or repel insects and animals that might otherwise consume them .
Periderm
In woody plants, the epidermis is sloughed off and replaced by a periderm after the cork cambium begins producing new tissues that increase the girth of the stem or root. The peri-derm constitutes the outer bark and is primarily composed of somewhat rectangular and boxlike cork cells, which are dead at maturity While the cytoplasm of cork cells is still functioning, it secretes a fatty substance, suberin, into the walls. This makes cork cells waterproof and helps them protect the phloem and other tissues beneath the bark from drying out, mechanical injury, and freezing temperatures. Some cork tissues, such as those produced by the cork oak, are harvested commercially and are used for bottle corks and in the manufacture of linoleum and gaskets. Some parts of a cork cambium form pockets of loosely arranged parenchyma cells that are not impregnated with suberin. These pockets of tissue protrude through the surface of the periderm; they are called and function in gas exchange between the air and the interior of the stem. The fissures in the bark of trees have lenticels at their bases. The various tissues discussed are shown as they occur in a woody stem in Figure 6.6.
Secretory Cells and Tissues
All cells secrete certain substances that can damage the cytoplasm, if allowed to accumulate internally. Such materials either must be isolated from the cytoplasm of the cells in which they originate or moved outside of the plant body. Often, the substances consist of waste products that are of no further use to the plant, but some substances, such as nectar, perfumes, and plant hormones, are vital to normal plant functions. Secretory cells may function individually or as part of a secretory tissue. Secretory cells or tissues, which often are derived from parenchyma, can occur in a wide variety of places in a plant. Among the most common secretory tissues are those that secrete nectar in flowers; oils in citrus, mint, and many other leaves; mucilage in the glandular hairs of sundews and other insect-trapping plants; latex in members of several plant families, such as the Spurge Family; and resins in coniferous plants, such as pine trees. Latex and resins are usually secreted by cells lining tubelike ducts that form networks throughout certain plant species Some plant secretions, such as pine resin, rubber, mint oil, and opium, have considerable commercial value.
Summary
A group of cells performing a common function is called a tissue. Apical meristems are found in the vicinity of the tips of roots and stems; the vascular cambium and the cork cambium occur as lengthwise cylinders within roots and stems; intercalary meristems occur in the vicinity of nodes of grasses and related plants.
Tissues produced by meristems consist of one to several kinds of cells. They include parenchyma, collenchyma, sclerenchyma, epidermis, xylem, phloem, periderm, and secretory tissues.
Parenchyma cells are thin-walled, while collenchyma cells have unevenly thickened walls that provide flexible support for various plant organs.
Two types of sclerenchyma occur—fibers (which are long and tapering) and sclereids (which are short in length); both types have thick walls and are usually dead at maturity.
Complex tissues have more than one kind of cell. The principal types are xylem, phloem, epidermis, and -periderm.
Xylem conducts water and minerals throughout the plant. It consists of a combination of parenchyma, fibers, vessels (tubular channels), tracheids (cells with tapering end walls that overlap), and ray cells (involved in lateral conduction).
Phloem conducts primarily dissolved sugars throughout the plant. It is composed of sieve tubes (made up of cells called sieve tube members), companion cells (that apparently regulate adjacent sieve tube members), arenchyma, ray cells, and fibers. Callose aids in plugging injured sieve tubes. Sieve cells, which have overlapping end walls, and adjacent albuminous
cells take the place of sieve tube members and companion cells in ferns and cone-bearing trees.
he epidermis is usually one cell thick, with fatty cutin (forming the cuticle) within and on the surface of the outer walls.
The epidermis may include guard cells that border pores called stomata; root hairs, which are tubular extensions of single cells; other hairs that consist of one to several cells; and glands that secrete protective substances.
Periderm, which consists of cork cells and loosely arranged groups of cells comprising lenticels involved in gas exchange,constitutes the outer bark of woody plants.
Secretory tissues occur in various places in plants; they secrete substances such as nectar, oils, mucilage, latex, and resins.