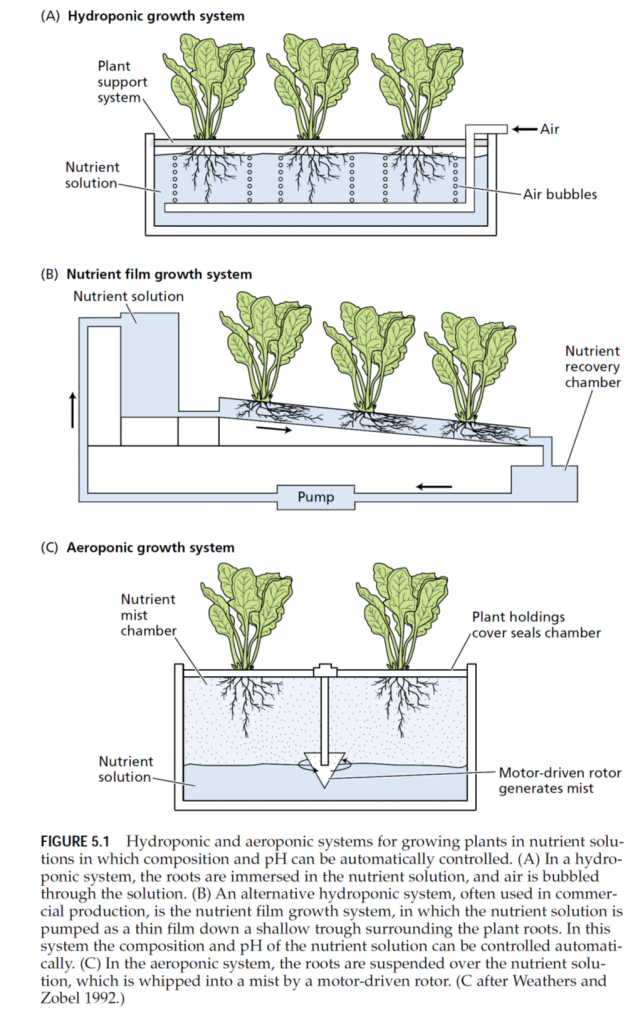
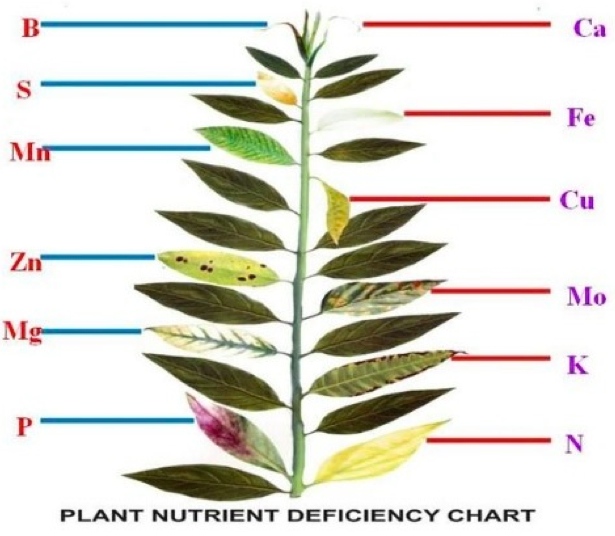
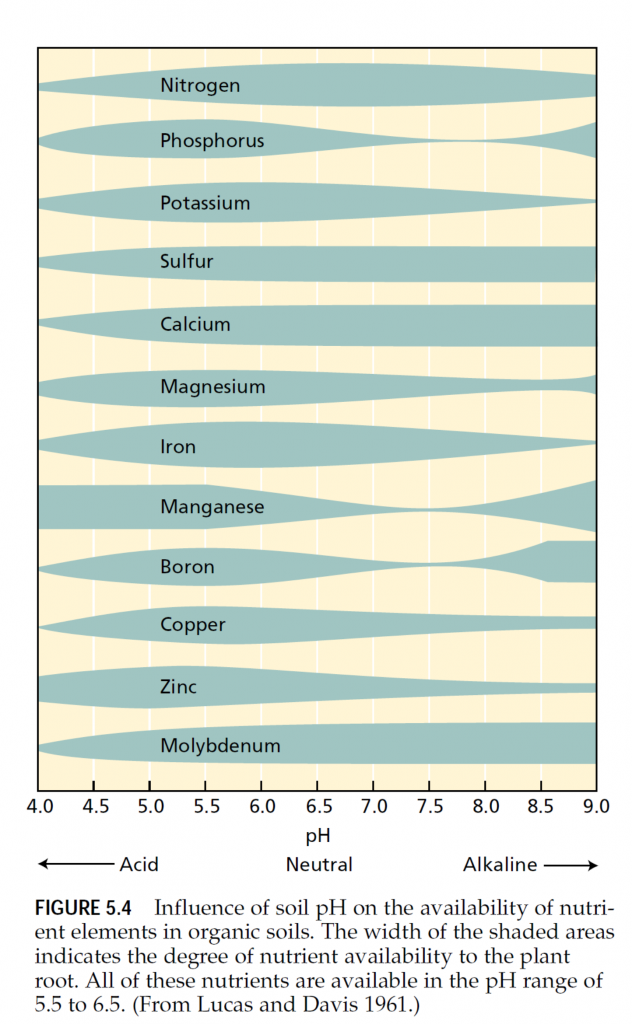
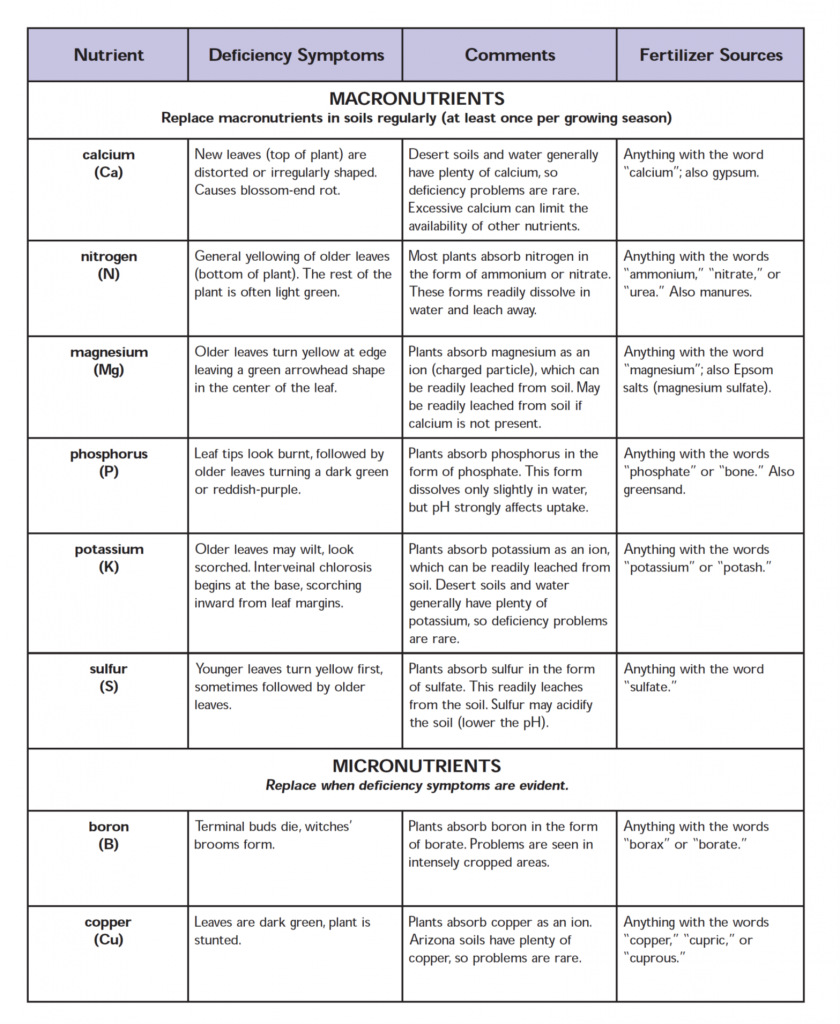
ENZYMES AND ENERGY TRANSFER
Enzymes (the proteins that speed up chemical reactions in cells without being used up in the reactions) regulate just about every
metabolic activity. In biochemical reactions, one or more specific enzymes are associated with the myriad forms of energy con- version that take place within cells. In some cases, these enzymes help build molecules that store energy in chemical bonds through a process called anabolism. Use of this energy to perform work often requires chemical bonds to be broken through catabolism. Most reactions of photosynthesis are anabolic because they involve construction of molecules that are stored for energy, whereas reactions in cellular respiration (referred to as “respiration” in the text) are generally catabolic, since in per- forming work, they release energy held in chemical bonds. Photosynthesis builds organic compounds by combining carbon di- oxide and water, forming carbohydrates. Respiration, on the other hand, breaks down those carbohydrates, producing carbon dioxide and water, which may be used once again in photosynthesis. As we will see in the sections to follow, this photosynthe- sis-respiration cycle is keyed by an enzyme complex that splits water molecules and releases electrons that function, at least temporarily, in storing biochemical energy. These electrons transfer energy from one form to another through oxidation- reduction reactions.
Introduction to metabolism: anabolism and catabolism | Khan Academy
Enzymes
Playlist-Enzymes/ Photosynthesis/respiration
Oxidation-Reduction Reactions
The processes of both photosynthesis and respiration include many oxidation-reduction reactions. Oxidation is the loss of one or more electrons; it involves removal of electrons from a compound. Reduction is the gain of one or more electrons; it involves the addition of electrons to a compound. In most oxidation-reduction reactions, oxidation of one compound is cou- pled with reduction of another compound catalyzed by the same enzyme or enzyme complex. When an electron is removed, a proton may follow, with the result that a hydrogen atom is often removed during oxidation and added during reduction. Oxy- gen is usually the oxidizing agent (i.e., the final acceptor of the electron), but oxidations can occur without oxygen being in- volved.
Krebs Cycle & Oxidative Phosphorylation
Krebs / citric acid cycle | Cellular respiration
Oxidation and reduction | Redox reactions and electrochemistry | Chemistry
Oxidation and reduction review from biological point-of-view | Biomolecules
PHOTOSYNTHESIS
The Essence of Photosynthesis
The energy-storing process of photosynthesis takes place in chloroplasts and other parts of green organisms in the pres- ence of light. The light energy stored in a simple sugar molecule is produced from carbon dioxide (CO2) present in the air and water (H2O) absorbed by the plant. When the carbon dioxide and water (H2O) are combined and ultimately a sugar molecule (C6H12O6, glucose) is produced in a chloroplast, oxygen gas (O2) is released as a by-product. The oxygen diffuses out into the atmosphere. The overall process can be depicted by the equation that follows. The equation should not, however, be taken literally because there are many intermediate steps to the process, and glucose is not the immediate first product of photosyn- thesis.
dioxide
6CO2 + 12H2O + light energy chlorophyll carbon water enzymes
C6H12O6 + 6O2 + 6H2O
¾¾¾¾®
glucose oxygen water
Photosynthesis takes place in chloroplasts (see Figs. 3.4 and 3.11) or in cells with membranes in which chlorophyll is embedded. The principals in the process are carbon dioxide, water, light, and chlorophyll; a brief examination of each fol- lows.
Carbon Dioxide
Water
Less than 1% of all the water absorbed by plants is used in photosynthesis; most of the remainder is transpired or incorporated into cytoplasm, vacuoles, and other materials. The water used is the source of electrons involved in photosynthesis, and the oxygen released is a by-product, even though carbon dioxide also contains oxygen. This has been demonstrated by conducting photosynthetic experiments using either carbon dioxide or water containing isotopes of oxygen. When the isotope is used only in the water, it appears in the oxygen gas released. If, however, it is used only in the carbon dioxide, it is confined to the sugar and water produced and never appears in the oxygen gas, demonstrating clearly that the water is the sole source of the oxygen released.
If water is in short supply, it may indirectly become a limiting factor in photosynthesis; under such circumstances, the stomata usually close and sharply reduce the carbon dioxide supply.
Light
Light that is too intense may change the way in which some of a cell’s metabolism takes place. For example, higher light intensities and temperatures may change the ratio of carbon dioxide to oxygen in the interiors of leaves, which, in turn, may accelerate photorespiration (discussed on p. 177). Photorespiration is typically considered to be a wasteful process that uses oxygen and releases carbon dioxide, although it may help some plants to survive under adverse conditions. It differs from common aerobic respiration in its chemical pathways.
Photooxidation, which involves the destruction (“bleaching”) of chlorophyll by light, may also occur. High light intensities may cause chlorophyll molecules to go to a different excited state. The energy released from the excited chlorophyll is passed to oxygen molecules, which become highly reactive and bleach the chlorophyll. In the fall, photooxidation plays a significant role in the breakdown of chlorophyll in leaves, resulting in the autumn colors discussed in Chapter 7. High light intensities may also cause an increase in transpiration, resulting in the closing of stomata. A sharp reduction in the available carbon dioxide supply inevitably follows.
Chlorophyll
There are several different types of chlorophyll molecules, all of which contain one atom of magnesium. They are very simi- lar in structure to the heme of hemoglobin, the iron–containing red pigment that transports oxygen in blood. Each molecule has a long lipid tail, which anchors the chlorophyll molecule in the lipid layers of the thylakoid membranes (Fig. 10.4).
Chloroplasts of most plants contain two major kinds of chlorophyll associated with the thylakoid membranes. Chloro- phyll a is blue-green in color and has the formula C55H72MgN4O5. Chlorophyll b is yellow-green in color and has the formula C55H70MgN4O6. Usually, a chloroplast has about three times more chlorophyll a than b. The more chlorophyll a there is in a cell, the brighter green the cell and the tissue of which it is a part appear to be. When a molecule of chlorophyll b absorbs light, it transfers the energy to a -molecule of chlorophyll a. Chlorophyll b, then, makes it possible for photosynthesis to take place over a broader spectrum of light than would be possible with chlorophyll a alone (see Fig. 10.7).
Other such pigments include carotenoids (yellowish to orange pigments), phycobilins (blue or red pigments found in cyanobacteria and red algae), and several other types of chlorophyll. Chlorophylls c, d, and possibly e take the place of chlo- rophyll b in certain algae, and several other photosynthetic pigments are found in bacteria. The various chlorophylls are all closely related and differ from one another only slightly in the structure of their molecules.
In chloroplasts, about 250 to 400 pigment molecules are grouped as a light-harvesting complex called a photosynthetic unit, with countless numbers of these units in each granum. Two types of these photosynthetic units function together in the chloroplasts of green plants, bringing about the first phase of photosynthesis, the light-dependent reactions, which are dis- cussed in the next section, “Introduction to the Major Steps of Photosynthesis.”
What are Electrons and Excitation?
Electrons and photons: absorption and transmission of light
Photoelectric effect | Electronic structure of atoms | Chemistry | Khan Academy
Leaf Pigments and Light
Photo-excitation of Chlorophyll
Introduction to the Major Steps of Photosynthesis
The process of photosynthesis takes place in two series of steps called the light-dependent reactions and the light- independent reactions. Although the light-independent reactions use products of the light-dependent reactions, both proc- esses occur simultaneously.
The Light-Dependent Reactions
The light-dependent reactions are the first major steps in the conversion of light energy to biochemical energy. The reactions are initiated when units of light energy (photons) strike chlorophyll molecules embedded in the thylakoid membranes of chloroplasts.
Our knowledge of the light-dependent reactions essentially began in the 1930s in England through a discovery by Robin Hill, a biochemist. He found that a solution of fragmented and whole chloroplasts, isolated from leaves that had been ground up and centrifuged, could briefly produce oxygen if an electron acceptor was present to receive electrons from water. In 1951, it was shown that NADP (nicotin-amide adenine dinucleotide phosphate), which is derived from the B vitamin niacin, was a natural electron acceptor in this reaction. In honor of its discoverer, the process became known as the Hill reaction.
During the light-dependent reactions,
- water molecules are split apart, producing electrons and hydrogen ions, and oxygen gas is released;
- the electrons from the split water molecules are passed along an electron transport system;
- energy-storing ATP molecules are produced;
- some hydrogen from the split water molecules is involved in the reduction of NADP to form NADPH (reduced nicoti- namide adenine dinucleotide phosphate), which carries hydrogen and is used in the second phase of photosynthesis, the light-independent reactions.
Conceptual overview of light dependent reactions
Light-Harvesting: The Antenna Complex
Photosynthesis Splitting H2O to O2 in Light Dependent Reactions 10 6 2018
Photosynthesis — View full playlist
Springlake-
plant videos 3 physiology
ps 1 – light harvesting
ps 2 – electron transport
ps 3 – reaction centers
The Light-Independent Reactions
The light-independent reactions (or carbon-fixing and reducing reactions) complete the conversion of light energy to chemi- cal energy in the form of ATP and NADPH. Some scientists refer to the light-independent reactions as the dark reactions because they don’t directly require light, but darkness has nothing to do with their functioning. In fact, even though light is not directly required in the same sense as it is for the light-dependent reactions, light is nevertheless required for the activa- tion of the enzymes involved, and the processes normally can occur only in daylight.
The light-independent reactions are a series of reactions that take place outside of the grana in the stroma of the chloro- plast (see page 40), if the products of the light-dependent reactions are available. They may initially proceed in different ways, depending on the particular kind of plant involved, but they all go through the Calvin cycle, discovered and elucidated by Dr. Melvin Calvin of the University of California. In 1961, Calvin received a Nobel Prize for unraveling how this most widespread type of light-independent reactions takes place.
In this cycle, carbon dioxide (CO2) from the air is combined with a 5-carbon sugar (RuBP, ribulose bisphosphate), and then the combined molecules are converted, through several steps, to sugars, such as glucose (C6H12O6). Energy and electrons involved in these steps are furnished by the ATP molecules and NADPH produced during the light-dependent reactions. Some of the sugars that are produced during the light-independent reactions are recycled, while others are stored as starch or
other polysaccharides (simple sugars strung together in chains). A summary of simplified photosynthetic reactions is shown in Figure 10.5. More detailed diagrams of the light-dependent and light-independent reactions are shown in Figures 10.8, 10.9, and 10.12.
Two molecules of a 3-carbon sugar compound (3PGA—an abbreviation of 3-phosphoglyceric acid) are shown as the first stable substance produced when carbon dioxide from the air and RuBP are combined and then converted during the light-independent reactions (Fig. 10.5). Some grasses and also many plants of arid regions fix carbon differently. They pro- duce a 4-carbon acid as the first product, followed by the Calvin cycle. This 4-carbon pathway is discussed, along with an- other variation found mostly in desert plants, in the next section, “A Closer Look at Photosynthesis.”
3. A Closer Look at Photosynthesis
A great deal has been learned about photosynthesis since 1772 when Joseph Priestly (1733–1804), a naturalist in England, re- ceived a medal for demonstrating that a sprig of mint “restored” oxygen so that a mouse could live in air that had been used up by a burning candle. Seven years later, Jan Ingen-Housz (1730–1799) of Holland, who visited England to treat the royal family for smallpox, carefully repeated Priestly’s demonstrations. He showed that the air was restored only when green parts of plants were receiving sunlight.
In 1782, Jean Senebier (1742–1809), a Swiss pastor, discovered that the photosynthetic process required carbon dioxide, and in 1796, Ingen-Housz showed that carbon went into the nutrition of the plant. The final component of the overall photo- synthetic reaction was explained in 1804 by another Swiss, Nicholas Theodore de Saussure (1767–1845), who showed that water was involved in the process.
A little current information about the details of photosynthesis is given in the following modest amplification of the pre- ceding section, “Introduction to the Major Steps of Photosynthesis.” Those who wish more information are referred to the reading list at the end of the chapter.
The Light-Dependent Reactions Reexamined
As noted earlier, light has characteristics of both waves and particles. Sir Isaac Newton, while experimenting with a prism over 300 years ago, produced a spectrum of colors from visible white light and postulated that light consisted of a series of discrete particles he called “corpuscles.”
Newton’s theory only partially explained light phenomena, however, and by the middle of the 19th century, James Max- well and others showed that light and all other parts of the extensive electromagnetic spectrum travel in waves.
By the late 1800s, with the discovery that a photoelectric effect can be produced in all metals, the wave theory also be- came inadequate to explain certain attributes of light. When a metal is exposed to radiation of a critical wavelength, it be- comes positively charged because the radiation forces electrons out of the metal atoms. The ability of light to force electrons from metal atoms depends on its particular wavelength—its energy content—and not on its intensity or brightness. The shorter the wavelength, the greater the energy, and vice versa.
In 1905, Albert Einstein proposed that the photoelectric effect results from discrete particles of light energy he called photons. In 1921, he received the Nobel Prize in physics for this work. Both waves and particles (photons) are today almost universally recognized as aspects of light. The energy (quantum) of a photon is not the same for all kinds of light; those of longer wavelengths have lower energy, and those of shorter wavelengths have proportionately higher energy.
Chlorophylls, the principal pigments of photosynthetic systems, absorb light primarily in the violet to blue and also in the red wavelengths; they reflect green wavelengths, which is why leaves appear green. This was first ingeniously demonstrated in 1882 by T. W. Engelmann. He focused a tiny spectrum of light on a filament (single row of cells) of Spirogyra, a freshwater alga. The alga had been mounted in a drop of water on a microscope slide containing bacteria that move toward an oxygen source. As shown in Fig. 10.6, the bacteria assembled in greatest numbers along the algal filament in the blue and red portions of the spectrum, demonstrating that oxygen production is directly related to the light the chlorophyll absorbs. An analysis using living material is called a bioassay. Information gained from bioassays often is significant.
Each pigment has its own distinctive pattern of light absorption, which is referred to as the pigment’s absorption spec- trum (Fig. 10.7). When a pigment absorbs light, the energy levels of some of the pigment’s electrons are raised. When this occurs, the energy may be emitted immediately as light (a phenomenon called fluorescence). In chlorophyll, the emitted light is characteristically in the red part of the visible light spectrum, so an extract of chlorophyll placed in light (especially ultra- violet or blue light) will appear red. The absorbed energy may also be emitted as light after a delay (a phenomenon called phosphorescence); it may otherwise be converted to heat. The most important use of the absorbed energy, however, is its storage in chemical bonds for photosynthesis.
Photosystems The two types of photosynthetic units present in most chloroplasts make up photosystems known as photo- system I and photosystem II (Fig. 10.8). Photosystem II received its “II” designation because it was discovered after photo- system I, but we know now that the events of photosynthesis that take place in photosystem II come before those of photosys- tem I.
While both photosystems produce ATP, only organisms that have photosystems I and II can produce NADPH and oxy-
gen as a consequence of electron flow. It is likely that evolutionary events led to organisms that possess both photosystems. At least 2.8 billion years ago, photosynthetic forms of bacteria (cyanobacteria) are believed to have evolved from primitive bacteria with the development of chlorophyll a and photosystem II. As a consequence of the oxygen-generating steps in pho- tosystem II that have evolved, humans and other organisms are dependent upon photosynthetic organisms that generate oxy- gen in the air we breathe today. Since photosynthetic organisms can generate oxygen from water, this very process in the future could be exploited to sustain human life during space travel and colonization of other planets.
Each photosynthetic unit of photosystem I consists of 200 or more molecules of chlorophyll a, small amounts of chloro- phyll b, carotenoid pigment with protein attached, and a special reaction-center molecule of chlorophyll called P700. Although all pigments in a photosystem can absorb photons, the reaction-center molecule is the only one that can actually use the light energy. The remaining photosystem pigments are called antenna pigments because together they function somewhat like an antenna in gathering and passing light energy to the reaction-center molecule (see Fig. 10.9). There are also iron-sulfur pro- teins that are the primary electron acceptors for photosystem I (i.e., iron-sulfur proteins first receive electrons from P700).
A photosynthetic unit of photosystem II consists of chlorophyll a, b-carotene (the precursor of vitamin A) attached to protein, a little chlorophyll b, and a reaction–center molecule of chlorophyll a; these special molecules are called P680.
The letter P stands for pigment and the numbers 700 and 680 of the reaction-center molecules of chlorophyll a refer to peaks in the absorption spectra of light with wavelengths of 700 and 680 nanometers, respectively. These peaks differ slightly from those of the otherwise identical chlorophyll a molecules of the photosynthetic units. A primary electron acceptor called pheophytin (or Pheo) is also present in photosystem II. One reaction-center molecule was found by Johann Deisenhofer and Hartmot Michel of Germany to have over 10,000 atoms. They received a Nobel Prize in 1988 for their work in determining the atomic structure.
Water-Splitting (Photolysis) When photons of light are absorbed by P680 molecules of a photosystem II reaction center (located near the inner surface of a thylakoid membrane), the light energy boosts electrons to a higher energy level. This is referred to as exciting electrons. Excited electrons are unstable and can rapidly revert back to their lower energy level, releasing absorbed en- ergy, perhaps as heat. However, during photosynthesis, the excited electrons are passed to an acceptor molecule, called pheo- phytin, at the beginning of an electron transport system.
From pheophytin, electrons are shuttled to another acceptor, PQ (plastoquinone), within the thylakoid membrane.4 PQ is mo- bile and moves through the lipid bilayer toward the inner side of the thylakoid membrane, unloading electrons to the cytochromes that are next in line. Electrons extracted from water by a manganese-containing oxygen-evolving complex (OEC) replace the elec- trons lost by the P680 molecule. It has been suggested that there is an oxidation-reduction system, usually designated Z, operating between water and P680. Transfer of electrons from Z to P680 reoxidizes Z and prepares it for accepting additional electrons from the OEC. Recent investigations indicate that for each two water molecules that are split, one molecule of oxygen is produced, along with four protons and four electrons.
This metabolic pathway eventually evolved in photosynthetic bacteria (cyanobacteria). The abundance of water, as an electron source, probably facilitated the mechanism that generated oxygen as a by-product of photosynthesis. This process increased the sup- ply of oxygen in earth’s atmosphere and made it possible for energy-efficient aerobic respiration to evolve.
Electron Flow and Photophospho-rylation The high-energy acceptor molecule PQ releases electrons that originated from photosystem II to an electron transport system, which functions something like a downhill bucket brigade. This electron transport system consists of iron-containing pigments called cytochromes and other electron transfer molecules, plus plastocyanin—a pro- tein that contains copper. When electrons pass along the electron transport system and protons subsequently move across the thy- lakoid membrane by chemiosmosis (see next section, “Chemiosmosis”), ATP molecules are formed from ADP in the process of – photophosphorylation.
A somewhat similar series of events occurs in photosystem I. When photons of light are absorbed by P700 molecules in a photo- synthetic unit, the energy excites electrons, which are transferred to an iron-sulfur acceptor molecule -designated Fe-S. This then passes electrons to another iron-sulfur acceptor molecule, Fd (ferredoxin), which, in turn, releases them to a carrier molecule desig- nated as FAD (flavin adenine dinucleotide). FAD contains flavoprotein, which assists in the reduction of NADP to NADPH. The electrons removed from the P700 molecule are replaced by electrons from photosystem II via the electron transport system outlined previously. This overall movement of electrons from water to photosystem II to photosystem I to NADP is called noncyclic electron flow, because it goes in one direction only. The production of ATP that correspondingly occurs is designated as noncyclic photophos- phorylation.
Photosystem I can also work independently of photosystem II. When it does, the electrons boosted from P700 reaction-center molecules (of photosystem I) are passed from ferredoxin to plastoquinone (instead of NADPH) and back into the reaction center of photosystem I. This process is cyclic electron flow. ATP generated by cyclic electron flow is called cyclic photophosphoryla- tion, but water molecules are not split and neither NADPH nor oxygen are produced in this process.
Chemiosmosis The oxygen-evolving complex on the inside of a thylakoid membrane catalyzes the splitting of water molecules, producing protons and electrons, as well as oxygen gas. These electrons used to replenish those excited in chlorophyll are then trans- ferred in bucket-brigade fashion through an electron transport system, ultimately reducing NADP to NADPH. As electrons travel through this transport system, additional protons move from the stroma to the inside of the thylakoid membrane in specific oxidation- reduction reactions when electrons pass from photosystem II to PQ. These protons join with the protons from the split water mole-
cules and thereby contribute to an accumulation of four protons toward the inside of the thylakoid membrane (an area also known as the thylakoid lumen).
Although some protons are used in the production of NADPH on the stroma side of the thylakoid membrane, there is still a net accumulation of protons in the thylakoid lumen from the splitting of water molecules and electron transport. This establishes a proton gradient, giving special proteins in the thylakiod membrane the potential for moving protons from the thylakoid lumen back to the stroma. Movement of protons across the membrane is thought to be a source of energy for the synthesis of ATP. The action has been described as similar to the movement of molecules during osmosis and has been called chemiosmosis, or the Mitchell theory, after its author, Peter Mitchell. In this concept, protons move across a thylakoid membrane through protein channels called ATPase. With the proton movement, ADP and phosphate (P) combine, producing ATP (Fig. 10.9). This chemiosmotic mechanism for connecting electron flow with conversion of ADP to ATP is essentially the same as that of oxida- tive phosphorylation in mitochondria (see Fig. 10.14), except that in mitochondria, oxygen (instead of NADP) is the terminal electron acceptor.
The Light-Independent Reactions Reexamined
We have seen how synthesis of ATP and NADPH is set in motion during the light-dependent reactions. Both of these substances play key roles in the synthesis of carbohydrate from carbon dioxide from the atmosphere, which during the light-independent re- actions reaches the interior of chlor-enchyma tissues via stomata. As indicated earlier, the light-independent reactions are really a whole series of reactions, each mediated by an enzyme in this major phase of photosynthesis.
The light-independent reactions take place in the stroma of the chloroplasts and, as long as the products of the light- dependent reactions are present, they do not directly need light in the same sense that the light-dependent reactions do. How- ever, they normally take place only during daylight hours because some of the enzymes involved in the light-independent reactions require light for their activation or conversion to a form in which they can actively catalyze steps of the light- independent reactions.
The Calvin Cycle The heart of the light-independent reactions is the Calvin cycle, during which carbon dioxide is fixed and converted to carbohydrate. The carbohydrate produced during these reactions facilitates growth, including the develop- ment of leaves, stems, roots, flowers, and other plant structures. From an ecological standpoint, this process is essential be- cause it is the main biosynthetic pathway through which carbon enters the web of life. As discussed next, the Calvin cycle, or photosynthetic carbon reduction (PCR) pathway (Fig. 10.10), runs in five main steps:
- Six molecules of carbon dioxide (CO2) from the air combine with six molecules of ribulose 1,5-bisphosphate (RuBP, the 5- carbon sugar continually being formed while photosynthesis is occurring), with the aid of the enzyme rubisco (RuBP car- boxylase/oxygenase).5
- The resulting six 6-carbon unstable complexes are immediately split into twelve 3-carbon molecules known as 3- phosphoglyceric acid (3PGA), the first stable compound formed in photosynthesis.
- The NADPH (which has been temporarily holding the hydrogen and electrons released during the light-dependent reac- tions) and ATP (also from the light-dependent reactions) supply energy and electrons that chemically reduce the 3PGA to twelve molecules of glyceraldehyde 3-phosphate (GA3P, 3-carbon sugar phosphate).
- Ten of the twelve glyceraldehyde 3-phosphate molecules are restructured, using another six ATPs, and become six 5- carbon molecules of RuBP, the sugar with which the Calvin cycle was initiated.
- This leaves a net gain of two GA3P molecules, which can contribute either to an increase in the carbohydrate content of the plant (glucose, starch, cellulose, or related substances) or can be used in pathways that lead to the net gain of lipids and amino acids.
Since rubisco catalyzes formation of the 3-carbon compound 3PGA as the first isolated product in these light– independent reactions, plants demonstrating this process are called C3 plants. However, as indicated by its name, the enzyme RuBP carboxylase/oxygenase has the potential to fix both CO2 through its carboxylase activity described by the Calvin cycle and O2 through its oxygenase activity. The oxygenase activity of rubisco makes C3 plants vulnerable to a process called photorespiration.
Photorespiration While the carboxylase activity of rubisco ultimately results in accumulation of carbohydrates, this same enzyme can exhibit an oxygenase activity that catalyzes combination of oxygen with RuBP and a subsequent pathway that releases carbon dioxide. This photorespiration may be interpreted as a wasteful process that competes with the carbon-fixing role of photosynthesis. However, the oxygenase activity of rubisco responsible for photorespiration is not necessarily an omi- nous alternative because it provides a salvage pathway to allow C3 plants to survive under hot, dry conditions. Evidence sug- gests that photorespiration helps to dissipate ATP and accumulated electrons from the light reactions, thereby preventing photooxidative damage.
Factors that determine the extent of this photorespiration in C3 plants include temperature, CO2:O2 ratio, and kinetic prop- erties of rubisco. Hot and dry climates generally promote photorespiration because stomata are closed under these conditions,
leading to a decreased CO2:O2 ratio within the leaf and near rubisco complexes. Oxygen gas can accumulate from photolysis in a leaf with closed stomata and photorespiration is more likely to occur, particularly when the carbon dioxide concentration drops roughly below 50 parts per million. When temperatures are cooler and moisture is present, the stomata are more likely to open, which leads to an increased CO2:O2 ratio. Under milder climatic conditions, C3 plants are more efficient at fixing carbon dioxide.
Photorespiration requires cooperation among chloroplasts, peroxisomes, and mitochondria to facilitate shuttling of in- termediates along the photorespiratory pathway. The products of photorespiration are the 2-carbon phosphoglycolic acid, which is processed to some extent in peroxisomes and eventually released as carbon dioxide in mitochondria, and the 3- carbon phosphoglyceric acid that can reenter the Calvin cycle. No ATP is produced by photorespiration.
The 4-Carbon Pathway Sugar cane, corn, sorghum, and at least 1,000 other species of tropical grasses or arid region plants sub- ject to high temperature stresses have a distinctive leaf -anatomy called Kranz anatomy (Fig. 10.11). Kranz anatomy leaves have two forms of chloroplasts. In the bundle sheath cells surrounding the veins, there are large chloroplasts, often with few to no grana. The large chloroplasts have numerous starch grains, and the grana, when present, are poorly developed. The chloroplasts of the mesophyll cells are much smaller, usually lack starch grains, and have well-developed grana.
Plants with Kranz anatomy produce a 4-carbon compound, oxaloacetic acid, instead of the 3-carbon 3–phosphoglyceric acid during the initial steps of the light-independent reactions. Oxaloacetic acid is produced when a 3-carbon compound, phosphoenolpyruvate (PEP), and carbon dioxide are combined in mesophyll cells with the aid of a different carbon-fixing enzyme, PEP carboxylase. Depending on the species, the oxaloacetic acid may then be converted to aspartic, malic, or other acids. PEP carboxylase is not sensitive to oxygen and, hence, has a greater enzyme affinity for carbon dioxide, so there is no photorespiratory loss of carbon dioxide captured in the organic acids. The carbon dioxide is transported to the bundle sheath cells where it is released and enters the normal Calvin cycle just as in C3 plants. The carbon dioxide concentration can be kept high in relation to the oxygen concentration in the bundle sheath cells, thereby keeping the reaction of rubisco with oxygen very low (Fig. 10.12).
Because the PEP system produces 4-carbon compounds, plants having this system are called 4-carbon, or C4, plants, to distinguish them from plants that have only the 3-carbon, or C3, system. Besides Kranz anatomy, C4 plants have other charac- teristic features:
- High concentrations of PEP carboxylase are found in the mesophyll cells. This is significant because PEP carboxylase allows the conversion of carbon dioxide to carbohydrate at much lower concentrations than does rubisco (found only in the bundle sheath cells), the corresponding enzyme in the Calvin cycle.
- The optimum temperatures for C4 photosynthesis are much higher than those for C3 photosynthesis, allowing C4 plants to thrive under conditions that would adversely affect C3 plants.
Obviously, the C4 pathway furnishes carbon dioxide to the Calvin cycle in a more roundabout way than does the C3 path- way, but the advantage of this extra pathway is a major reduction of photorespiration in C4 plants. Also, in C4 plants, the C4 pathway in the mesophyll cells results in carbon dioxide being picked up even at low concentrations (via the enzyme PEP car- boxylase) and in carbon dioxide being concentrated in the bundle sheath, where the Calvin cycle takes place almost exclusively. The Calvin cycle enzyme, rubisco, will catalyze the reaction in which RuBP will react with carbon dioxide rather than oxygen. Consequently, C4 plants, which typically photosynthesize at higher temperatures, have photosynthetic rates that are two to three times higher than those of C3 plants. However, at lower temperatures, C3 photosynthesis is more efficient than that of C4 plants because the cost of photorespiration at those temperatures is usually less than the two extra ATPs required for the C4 pathway.
CAM Photosynthesis Crassulacean acid metabolism (CAM) photosynthesis is found in plants of about 30 families, includ- ing cacti, stonecrops, orchids, bromeliads, and other succulents that are often stressed by limited availability of water. A few succulents do not have CAM photosynthesis, however, and several nonsucculent plants do. Many CAM plants are facultative C3 plants that can switch to C3 photosynthesis during the day after a good rain or when night temperatures are high. Plants with CAM photosynthesis typically do not have a well-defined palisade mesophyll in the leaves, and, in contrast to the chloroplasts of the bundle sheath cells of C4 plants, those of CAM photosynthesis plants resemble the mesophyll cell chloro- plasts of C3 plants.
CAM photosynthesis is similar to C4 photosynthesis in that 4-carbon compounds are produced during the light–independent reactions. In these plants, however, the organic acids (mainly malic acid) accumulate at night and break down during the day, re- leasing carbon dioxide. The enzyme PEP carboxylase is responsible for converting the carbon dioxide plus PEP to organic acids at night when the stomata are open. During daylight, the organic acids diffuse out of the cell vacuoles in which they are stored and are converted back to carbon dioxide for use in the Calvin cycle. A much larger amount of carbon dioxide can be converted to carbohydrate each day than would otherwise be possible, since the stomata of such plants are closed during the day to conserve water. This arrangement allows the plants to function well under conditions of both limited water supply and high light intensity (Fig. 10.13).
Other Significant Processes That Occur in Chloroplasts
In addition to photosynthesis, there are two very important sets of biochemical reactions that take place in chloroplasts (and also in the proplastids of roots).
- Sulfates are reduced to sulfide via several steps involving ATP and enzymes. The sulfide is rapidly converted into impor- tant sulfur-containing amino acids, such as methionine and cysteine, which are part of the building blocks for proteins, anthocyanin pigments, chlorophylls, and several other cellular components.
- Nitrates are reduced to organic compounds. Initially, the nitrates are reduced to nitrites in the cytoplasm. The nitrites are then transported into chloroplasts (or root proplastids) where, through several enzyme-mediated steps, they are converted to ammonia. The ammonia is then converted to amino groups that are integral parts of several important amino acids such as glutamine and aspartic acid. Glutamine is an important form of nitrogen storage in roots or specialized stems such as those of carrots, beets, and potato tubers.
RESPIRATION
The solar energy that is converted into biochemical energy by the process of photosynthesis today is stored in various organic compounds such as wood, while coal and oil contain energy originally captured by green organisms in the geological past. If the organic compounds are burned, the energy is released very rapidly in the form of heat and light, and much of the usable energy is lost. Living organisms, however, “burn” their energy-containing compounds in numerous, small, enzyme-controlled steps that release tiny amounts of immediately usable energy. The released energy is usually stored in ATP molecules, which allows the available energy to be used more efficiently and the process to be controlled more precisely.
The Essence of Respiration
Respiration is essentially the release of energy from glucose molecules that are broken down to individual carbon dioxide molecules. The process takes place in all active cells 24 hours a day, regardless of whether or not photosynthesis happens to be occurring simultaneously in the same cells. It is initiated in the cytoplasm and completed in the mitochondria. The energy, stored in chemical bonds containing high-energy electrons, is released from simple sugar molecules that are broken down during a series of steps controlled by enzymes. No oxygen is needed to initiate the process, but in aerobic respiration (the most widespread form of respiration), the process cannot be completed without oxygen gas (O2). The controlled release of energy is the significant event; carbon dioxide (CO2) and water (H2O) are the by-products. Aerobic respiration can be summed up in the following equation, but bear in mind that respiration, like photosynthesis, is a complex process that in- volves many steps not reflected in a simplified equation.
C6H12O6 + 6O2 enzymes 6CO2 + 6H2O + energy glucose oxygen ¾¾® carbon water
dioxide
Anaerobic respiration and fermentation are two forms of respiration that were probably carried on in the geological past when there was no oxygen in the atmosphere. These forms of respiration are still carried on today by certain bacteria and other organisms in the absence of oxygen gas. Anaerobic respiration and fermentation release less than 6% of the energy re- leased from a molecule of glucose by aerobic respiration. The two forms differ from one another in the manner in which hy- drogen released from the glucose is combined with other substances (see the discussion on p. 183). Fermentation is very im- portant industrially, particularly in the brewing and baking industries. Two well-known forms of fermentation are illustrated by the following equations:
C6H12O6 enzymes 2C2H5OH + 2CO2 + energy (ATP) glucose õõõã ethyl carbon
alcohol dioxide
C6H12O6 enzymes 2C3H6O3 + energy (ATP)
glucose õõõã lactic acid
The relatively small amount of energy released during these inefficient forms of respiration is partly stored in two ATP molecules. The actual amount of energy stored is roughly only 29% of the approximately 48 Kcals6 of energy released in anaerobic respiration.
Introduction to the Major Steps of Respiration
Glycolysis
In most forms of carbohydrate respiration, the first major phase takes place in the cytoplasm and requires no oxygen gas (O2). This phase, called glycolysis, involves three main steps and several smaller ones, each controlled by an enzyme. During the process, a small amount of energy is released, and some hydrogen atoms are removed from compounds derived from a glu- cose molecule. The essence of this complex series of steps is as follows:
- In a series of reactions, the glucose molecule becomes a fructose molecule carrying two phosphates (P).
- This sugar (fructose) molecule is split into two 3-carbon fragments called glyceraldehyde 3-phosphate (GA3P).
- Some hydrogen, energy, and water are removed from these 3-carbon fragments, leaving pyruvic acid (Fig. 10.14).
Two ATP molecules supply the energy needed to start the process of glycolysis. By the time pyruvic acid has been formed, however, four ATP molecules have been produced from the energy released along the way, for a net gain of two ATP molecules. A great deal of the energy originally in the glucose molecule remains in the pyruvic acid. The hydrogen ions and high-energy electrons released during the process are picked up and temporarily held by an acceptor molecule, NAD (nicotinamide adenine dinucleotide). What happens to them next depends on the kind of respiration involved: aerobic respi- ration, true anaerobic respiration, or fermentation.
Aerobic Respiration
In aerobic respiration (the most common type of respiration), glycolysis is followed by two major stages: the citric acid cycle and electron transport. Both stages occur in the mitochondria and involve many smaller steps, each of which is controlled by enzymes (see Fig. 10.14).
The Citric Acid (Krebs) Cycle The citric acid cycle was originally named the Krebs cycle after Hans Krebs, a British biochemist who received a Nobel Prize in 1953 for his unraveling of many of the complex reactions that take place in respira- tion. The name citric acid cycle, or tricarboxylic acid (TCA) cycle, reflects the important role played by several organic acids during the process.
Before entering the citric acid cycle, which takes place in the fluid matrix located within the compartments formed by the cristae of mitochondria (see Fig. 3.13), carbon dioxide is released from pyruvic acid that was produced by glycolysis. What remains is restructured to a 2-carbon acetyl group. This acetyl group combines with an acceptor molecule called coen- zyme A (CoA). This combination (called acetyl CoA) then enters the citric acid cycle, which is a series of biochemical reac- tions that are catalyzed by enzymes. Little of the energy originally trapped in the glucose molecule is released during glyco- lysis. As the citric acid cycle proceeds, however, high-energy electrons and hydrogen are successively removed. This re- moval takes place from a series of organic acids and, after transfer, ultimately produces compounds such as NADH (re- duced nicotinamide adenine dinucleotide) and FADH2 (reduced flavin adenine dinucleotide), as well as a small amount of ATP. Carbon dioxide is produced as a by-product while the cycle is proceeding.
Electron Transport Much of the energy originally in the glucose molecule now has been transferred to the acceptors NAD and FAD, which became NADH and FADH2, respectively. NADH and FADH2 are electron donors to an electron transport system consisting of special acceptor molecules arranged in a precise sequence on the inner membrane of mitochondria. The electrons flow through a series of carrier molecules, many of which are part of protein complexes, down an energy gradient. Some of these electron carriers also accept protons and release them to the intermembrane space of the mitochondrion. Shut- tling of protons in this way causes a build-up of protons outside the mitochondrial matrix, thereby establishing an electro- chemical gradient. Through the process of chemiosmosis, additional protein complexes couple the transport of protons back into the matrix with phosphorylation of ADP to form ATP. The production of ATP stops if there are no electron donors or electron acceptor oxygen.
The acceptor molecules include iron-containing proteins called cytochromes. Energy is released in small increments at each step along the system, and ATP is produced from ADP and P. As the final step in aerobic respiration (see Fig. 10.14), oxygen acts as the ultimate electron acceptor, producing water as it combines with hydrogen.
By the time the process is complete, the recoverable energy locked in a molecule of glucose has been released and is stored in ATP molecules. This stored energy is then available for use in the synthesis of other molecules and for growth, active transport, and a host of other metabolic processes. Aerobic respiration produces a net gain of 36 ATP molecules from one glucose molecule, using up six molecules of oxygen and producing six molecules of carbon dioxide and a net total of six molecules of water. For each mole (180 grams) of glucose aerobically respired, 686 Kcal of energy is re- leased, with about 39% of it being stored in ATP molecules and the remainder being released as heat.
Anaerobic Respiration and Fermentation
In living organisms, glucose molecules often may undergo glycolysis without enough oxygen being available to complete
aerobic respiration. In such cases, the hydrogen released during glycolysis is simply transferred from the hydrogen acceptor molecules back to the pyruvic acid after it has been formed, creating ethyl alcohol in some organisms, and lactic acid or simi- lar substances in others. A little energy is released during either fermentation or true anaerobic respiration, but most of it re- mains locked up in the alcohol, lactic acid, or other compounds produced.
In true anaerobic respiration, the hydrogen removed from the glucose molecule during glycolysis is combined with an inorganic ion, as, for example, when sulfur bacteria (discussed in Chapter 17) convert sulfate (SO4) to sulfur (S) or another sulfur compound or when certain cellulose bacteria produce methane gas (CH4) by combining the hydrogen with carbon di- oxide.
Oxygen gas is not required to make these compounds, but few organisms can live long without oxygen, and many that carry on fermentation can also respire aerobically. If oxygen becomes available, the remaining energy can be released by further breakdown of these compounds. About 7% of the total energy in a glucose molecule is removed during anaerobic respiration or fermentation. So much of that energy goes into the making of the alcohol or the lactic acid or is dissipated as heat that there is a net gain of only two ATP molecules (compared with 36 ATP molecules produced in aerobic respiration). The forms of anaerobic respiration are adaptive to the organisms that have them in that they recycle NAD and allow glycoly- sis to continue.
Living cells can tolerate only certain concentrations of alcohol. In media in which yeasts are fermenting sugars, for exam- ple, once the alcohol concentration builds up beyond 12%, the cells die and fermentation ceases. This is why most wines have an alcohol concentration of about 12% (24 proof).
Many bacteria carry on both fermentation and true anaerobic respiration simultaneously, making it difficult to distinguish between the two processes. Some texts use the terms anaerobic respiration and fermentation interchangeably to designate respiration occurring in the presence of little or no oxygen gas.
Factors Affecting the Rate of Respiration
Temperature
Temperature plays a major role in the rate at which the various respiratory reactions occur. For example, when air tempera- tures rise from 20°C (68°F) to 30°C (86°F), the respiration rates of plants double and sometimes even triple. The faster respi- ration occurs, the faster the energy is released from sugar molecules, with an accompanying decrease in weight. In growing plants, this weight loss is more than offset by the production of new sugar by photosynthesis. In harvested fruits, seeds, and vegetables, however, respiration continues without sugar replacement, and some water loss also occurs. Respiring cells con- vert energy stored as starch or sugar primarily to ATP, but much of the energy is lost in the form of heat, with only 39% be- ing stored as ATP. Most fresh foods are kept under refrigeration, not only to lower the respiration rate and retard water loss, but also to dissipate the heat. Keeping the temperatures down is also important to prevent the growth and reproduction of food-spoiling molds and bacteria, which may thrive at warmer temperatures.
Heat inactivates most enzymes at temperatures above 40°C (104°F), but a few organisms, such as various cyanobacteria and algae in the hot springs of Yellowstone National Park and similar places, have adapted in such a way that they are able to thrive at temperatures exceeding 60°C (140°F)—heat that would kill other organisms of comparable size almost instantly.
Water
Water inside the cells and their organelles act as a medium in which the enzymatic reactions can take place. Living cells often have a water content of more than 90%, but the cells of mature seeds may have a water content of less than 10%. When water content becomes this low, respiration does not cease completely, but it continues at a drastically reduced rate, resulting in only very tiny amounts of heat being released and of carbon dioxide being given off. Seeds may remain viable (capable of germinat- ing) for many years if stored under dry conditions. If they come in contact with water, however, they swell by imbibition. Respi- ration rates then increase rapidly. If the wet seeds happen to be in an unrefrigerated storage bin, the temperature may increase to the point of killing the seeds. In fact, if fungi and bacteria begin to grow on the seeds, temperatures from their respiration can become so high that spontaneous combustion can sometimes occur.
Oxygen
If flooding sharply reduces the oxygen supply available to the roots of trees and house plants, their respiration and growth rates may be decreased. They may even die if the condition persists too long. When foods are stored, however, it helps to bring about lower rates of respiration by reducing the oxygen in the storage areas. In fact, it is a common commercial practice to reduce the oxygen present in warehouses where crops are stored. The oxygen content is reduced to as little as 1% to 3% by pumping in nitrogen gas, while maintaining low temperatures and humidity. Oxygen concentration is not reduced below 1% because that can result in an undesirable increase in fermentation.
3. A Closer Look at Respiration
Respiration, like photosynthesis, is a very complex process, and, as with photosynthesis, it is beyond the scope of this book to explore the subject in great detail. The following amplification of information already discussed is modest, and those who wish further information are referred to the reading list at the end of the chapter.
Glycolysis Reexamined
As previously discussed, this initial phase of all forms of respiration brings about the conversion of each 6-carbon glucose molecule to two 3-carbon pyruvic acid molecules via three main steps, each mediated by enzymes. The three main steps are as follows:
- Phosphorylation, whereby the 6-carbon sugars receive phosphates
- Sugar cleavage, which involves the splitting of 6–carbon fructose into two 3-carbon sugar fragments
- Pyruvic acid formation, which involves the oxidation of the sugar fragments
Energy needed to initiate the process is furnished by an ATP molecule, which also furnishes the phosphate group for the phos- phorylation of the sugar glucose to yield glucose 6-phosphate. Another ATP, with the aid of the enzyme fructokinase, yields fruc- tose bisphosphate (fructose 1,6-diphosphate). As a result of the cleavage of the fructose bisphosphate, two different 3-carbon sugars are produced, but ultimately, only two glyceraldehyde 3-phosphate (GA3P) molecules remain. These two 3-carbon sugars are oxi- dized to two 3-carbon acids, and, in the successive production of several of these acids, phosphate groups are removed from the acids. The phosphate groups combine with ADP, producing a net direct gain of two ATP molecules during glycolysis. In addition, hydrogen is removed as GA3P is oxidized. This hydrogen is picked up by the acceptor molecule, NAD, which becomes NADH. Glycolysis, which requires no oxygen gas, is summarized in Figure 10.14.
Transition Step to the Citric Acid (Krebs) Cycle
Before a pyruvic acid molecule enters the citric acid cycle, which takes place in the mitochondria, a molecule of carbon dioxide is removed and a molecule of NADH is produced, leaving an acetyl fragment. The 2-carbon fragment is then bonded to a large mole- cule called coenzyme A. Coenzyme A consists of a combination of the B vitamin pantothenic acid and a nucleotide. Pantothenic acid is one of several B vitamins essential to respiration in both plants and animals; others include thiamine (vitamin B1), niacin, and riboflavin. The bonded acetyl fragment and coenzyme A molecule is referred to as acetyl CoA. The following equation summarizes the fate of the two pyruvic acid molecules following glycolysis and leading to the citric acid cycle:
2 pyruvic acid + 2 CoA + 2 NAD õõõõã 2 acetyl CoA + 2 NADH + 2CO2
In addition to pyruvic acid, fats and amino acids can also be converted to acetyl CoA and enter the process at this point. The NADH molecules donate their hydrogen to an electron transport system (discussed in the section, “Electron Transport and Oxidative Phosphorylation”), and the acetyl CoA enters the citric acid cycle (see Fig. 10.14).
The Citric Acid (Krebs) Cycle Reexamined
In the citric acid cycle, acetyl CoA is first combined with oxaloacetic acid, a 4-carbon compound, producing citric acid, a 6- carbon compound. The citric acid cycle is kept going by oxaloacetic acid, which is produced in small amounts, but is an inter- mediate product rather than a starting substance or an end product of the cycle. As the cycle progresses, a carbon dioxide is removed, producing a 5-carbon compound. Then another carbon dioxide is removed, producing a 4-carbon compound. This 4- carbon compound, through additional steps, is converted back to oxaloacetic acid, the substance with which the cycle began, and the cycle is repeated.
Each full cycle uses up a 2-carbon acetyl group and releases two carbon dioxide molecules while regenerating an ox- aloacetic acid molecule for the next turn of the cycle. Some hydrogen is removed during the process and is picked up by FAD and NAD. One molecule of ATP, three molecules of NADH, and one molecule of FADH2 are produced for each turn of the cycle. The citric acid cycle may be summarized as follows:
oxaloacetic + acetyl + ADP + P + 3 NAD + FAD õõõõã acid CoA
oxaloacetic + CoA + ATP + 3 NADH + H+ + FADH2 + 2CO2 acid
The hydrogen carried by NAD and FAD can mostly be traced to the acetyl groups and to water molecules added to some compounds in the citric acid cycle. The FAD and FADH2 are now known to be intermediate compounds. Ubiquinol, a com- ponent of the electron transport system, receives electrons from either NADH or FADH2.
Electron Transport and Oxidative Phosphorylation
After completion of the citric acid cycle, the glucose molecule has been totally dismantled, and some of its energy has been transferred to ATP molecules. A considerable portion of the energy was transferred to NAD and FAD when they were used to pick up hydrogen and electrons from the molecules derived from glucose as they were broken down during glycolysis and the citric acid cycle. This energy is released as the hydrogen and electrons are passed along an electron transport system. This sys- tem, like the electron transport system of photosynthesis, functions something like a high-speed bucket brigade in passing along electrons from their source to their destination. Several of the electron carriers in the transport system are cytochromes. They are very specific and, as electrons flow along the system, they can transfer their electrons only to other specific acceptors. When the electrons reach the end of the system, they are picked up by oxygen and combine with hydrogen ions, forming water.
Part of the energy that is released during the movement of electrons along the electron transport system can be used to make ATP in a process called oxidative phosphorylation. If hydrogen ion and electron transport begins with NADH, which was produced inside the mitochondria (i.e., during the conversion of pyruvic acid to acetyl CoA and during the citric acid cycle), enough energy is produced to yield three ATP molecules from each NADH molecule. Similarly, if hydrogen ion and electron transport begins either with FADH2 or with NADH, produced outside the mitochondria (i.e., during glycolysis), two ATP molecules are produced.
The manner in which ATP is produced during the operation of the respiratory electron transport system involves essen- tially the same chemiosmotic concept that was applied earlier to proton movement across thylakoid membranes.
The chemiosmosis theory concerning electron transport and proton movement across membranes was proposed in the 1960s by Peter Mitchell, a British biochemist, and is now widely accepted as the explanation for the movements in both pho- tosynthesis and respiration. In respiration, oxidative phosphorylation is energized by a gradient of protons (H+) that flow by chemiosmosis across the inner membrane of a mitochondrion. Mitchell, who received a Nobel Prize for his work in 1978, surmised that protons are “pumped” from the matrix of the mitochondria to the region between the two membranes (see Fig. 3.13) as electrons flow from their source in NADH molecules along the electron transport system, which is located in the inner membrane. The protons are believed to “diffuse” back into the matrix via channels provided by an enzyme complex known as the F1 particle (an ATPase), releasing energy that is used to synthesize ATP.
If we retrace our steps through the entire process of aerobic respiration, we find that glycolysis yields four -molecules of ATP and two molecules of NADH (from which more molecules of ATP are formed), for a total of eight ATP from the con- version of glucose to two pyruvic acid molecules. Two ATP are used in the process, however, leaving a net gain of six ATP.
When two pyruvic acid molecules are converted to two acetyl CoA in the mitochondria, two more NADH molecules (which will generate six molecules of ATP) are produced. The two acetyl CoA molecules metabolized in the citric acid cycle yield two molecules of ATP, two molecules of FADH2 (from which four ATP are formed), and six molecules of NADH (which cause the formation of 18 molecules of ATP), making a citric acid cycle total of 24 ATP. A grand total of 36 ATP is produced for the aerobic respiration of one glucose molecule (Table 10.1). The 36 ATP molecules represent about 39% of the energy originally present in the glucose molecule. The remaining energy is lost as heat or is unavailable. Aerobic respiration is still about 18 times more efficient than anaerobic respiration.
A condensed comparison between photosynthesis and respiration is shown in Table 10.2.
ADDITIONAL METABOLIC PATHWAYS
While photosynthesis and respiration are the main processes through which plants grow, develop, reproduce, and survive, there are many additional processes that contribute toward these activities. Most of these use intermediate steps, but they could not function without photosynthesis and respiration. Some of the essential compounds produced from additional path- ways include sugar phosphates and nucleotides, nucleic acids, amino acids, proteins, chlorophylls, cytochromes, carotenoids, fatty acids, oils, and waxes.
Metabolic processes not required for normal growth and development are generally referred to as secondary metabolism. Although not essential, many of the products from secondary metabolism enable plants to survive and persist under special conditions. These products provide the plant with unique colors, aromas, poisons, and other compounds that may attract or deter other organisms or give them a competitive edge in nature. Humans have exploited many secondary compounds from plants for medicinal, culinary, or other purposes. It has been estimated that 50,000 to 100,000 such compounds exist in plants with only a few thousand of these thus far having been identified. Secondary metabolic products may be derived from modi- fication of amino acids and related compounds to produce alkaloids or through specialized conversions such as the shikimic acid pathway (phenolics) and mevalonic acid pathway (terpenoids). Examples of these compounds are shown in Table 10.3. Lignin, which is a component of secondary cell walls, is, for example, synthesized through the shikimic acid pathway. Be- cause it is hard to digest and is toxic to some predators, it protects plants from herbivorous animals.
ASSIMILATION AND DIGESTION
Sugars produced through photosynthesis may undergo many transformations. Some sugars are used directly in respiration, but others not needed for that purpose may be transformed into lipids, proteins, or other carbohydrates. Among the most im- portant carbohydrates produced from simple sugars are sucrose, starch, and cellulose. Much of the organic matter produced through photosynthesis is eventually used in the building of protoplasm and cell walls. This conversion process is called as- similation.
When photosynthesis is taking place, sugar may be produced faster than it can be used or transported away to other parts of the plant. When this happens, the excess sugar may be converted to large, insoluble molecules, such as starch or oils, tem- porarily stored in the chloroplasts and then later changed back to a soluble form that is transported to other cells. The conver- sion of starch and other insoluble carbohydrates to soluble forms is called digestion (Fig. 10.15). The process is nearly al- ways one of hydrolysis, in which water is taken up and, with the aid of enzymes, the links of the chains of simple sugars that comprise the molecules of starch and similar carbohydrates are broken by the addition of water. The disaccharide malt sugar (maltose), for example, is transformed to two molecules of glucose, with the aid of an enzyme (maltase), by the addition of one molecule of water, as follows:
¾¾¾®
C12H22O11 + H2O maltase 2C6H12O6
maltose water (enzyme) glucose
Fats are broken down to their component fatty acids and glycerol, and proteins are digested to their amino acid building blocks in similar fashion. Digestion is carried on in any cell where there may be stored food, with very little energy being released in the process. In animals, special digestive organs also play a role in digestion, but plants have no such additional “help” in the process. In both plants and animals, digestion within cells is similar and is a normal part of metabolism.
Except in insect-trapping plants, digestion takes place within plant cells where the carbohydrates, fats, or proteins are stored, while in animals, digestion usually occurs outside of the cells in the digestive tract. Apart from the location, the proc- ess is essentially similar in plants and animals.
Summary
- Enzymes catalyze reactions of metabolism. Many of these include oxidation-reduction reactions. Oxidation is loss of electrons; reduction is gain of electrons.
- Photosynthesis is an anabolic process that combines carbon dioxide and water in the presence of light with the aid of chlo- rophyll; oxygen is a by-product. All life depends on photosynthesis, which takes place in chloroplasts.
- Carbon dioxide constitutes 0.037% of the atmosphere, but the percentage has been rising in recent years. Increased car- bon dioxide levels have potential to elevate global temperatures through the “greenhouse effect.”
- Chlorophyll b and carotenoids are antenna pigments that direct light energy to chlorophyll a. Photosynthetic units con- taining chlorophylls and accessory pigments absorb units of light energy, become excited, and pass this energy to accep- tors during the light-dependent reactions of photosynthesis.
- During the light-dependent reactions of photosynthesis, which occur in thylakoid membranes of chloroplasts, water molecules are split, and oxygen gas is released. Hydrogen ions and electrons are released from water and transferred to produce NADPH and ATP.
- The two types of photosynthetic units present in most chloroplasts are photosystems I and II. The events that take place in photosystem II come before those of photosystem I. Each photosystem has a reaction-center molecule of chlorophyll a that boosts electrons to a higher energy level when it is excited by light energy.
- Photosystem II boosts electron excitation to a level that, when it encounters photosystem I, has the potential to reduce NADP to NADPH through noncyclic electron flow. Photosystem I, by itself, can cycle electrons for generation of ATP. Electron transport while the photosystems are operating and proton movement across thylakoid membranes are both in- volved in ATP production.
- The light-independent reactions occur through a series of reactions known as the Calvin cycle, which takes place in the stroma of chloroplasts. In the first step, carbon dioxide is combined with RuBP through catalytic action of the enzyme rubisco to form two molecules of the 3-carbon compound, GA3P. The ATP and NADPH from the light-dependent reac- tions furnish energy to eventually convert GA3P to 6-carbon carbohydrates. This cycle also regenerates RuBP to enable continued carbon fixation.
- In the light-independent reactions of C4 plants, 4-carbon oxaloacetic acid is initially produced instead of 3-carbon PGA. In the leaf mesophyll of C4 plants, there are large chloroplasts, which contain rubisco in the bundle sheaths, and small
chloroplasts, which contain higher concentrations of PEP carboxylase that facilitate the conversion of carbon dioxide to carbohydrate at much lower concentrations than is possible in C3 plants.
- CAM photosynthesis occurs in succulent plants whose stomata are closed and admit little CO2 during the day. Regular photosynthesis occurs as the 4-carbon compounds that accumulate at night are converted back to carbon dioxide during the day.
- Respiration is a catabolic process that takes place in the cytoplasm and mitochondria of cells. The energy is released, with the aid of enzymes, from simple sugar and organic acid molecules.
- In aerobic respiration, stored energy release requires oxygen; CO2 and water are by-products of the process.
- Anaerobic respiration and fermentation do not require oxygen gas, and much less energy is released. The remaining energy is in the ethyl alcohol, lactic acid, or other such substances produced. Some released energy is stored in ATP molecules. Temperature, available water, and environmental oxygen affect respiration rates.
- Glycolysis, which occurs in the cytoplasm, requires no molecular oxygen; two phosphates are added to a 6-carbon sugar molecule, and the prepared molecule is split into two 3-carbon sugars (GA3P). Some hydrogen, energy, and water are removed from the GA3P, producing pyruvic acid. There is a net gain of two ATP molecules. Hydrogen ions and elec- trons released during glycolysis are picked up by NAD, which becomes NADH.
- In aerobic respiration, which occurs in the mitochondria, pyruvic acid loses some CO2, is restructured, and becomes ace- tyl CoA. Energy, CO2, and hydrogen are removed from the acetyl CoA in the citric acid cycle, which involves enzyme- catalyzed reactions of a series of organic acids.
- NADH passes the hydrogen gained during glycolysis and the citric acid cycle along an electron transport system; small increments of energy are released and partially stored in ATP molecules, and the hydrogen is combined with oxygen gas, forming water in the final step of aerobic respiration.
- Hydrogen removed from glucose during glycolysis is combined with an inorganic ion in anaerobic respiration. The hy- drogen is combined with the pyruvic acid or one of its derivatives in fermentation. Both processes occur in the absence of oxygen gas, with only about 7% of the total energy in the glucose molecule being released, for a net gain of two ATP molecules.
- Two molecules of NADH and two ATP molecules are gained during glycolysis when two 3-carbon pyruvic acid mole- cules are produced from a single glucose molecule. Another molecule of NADH is produced when the pyruvic acid molecule is restructured and becomes acetyl CoA prior to entry into the citric acid cycle.
- In the citric acid cycle, acetyl CoA combines with 4-carbon oxaloacetic acid, producing first a 6-carbon compound, next a 5-carbon compound, and then several 4-carbon compounds. The last 4-carbon compound is oxaloacetic acid. Two CO2 molecules are also released during this process.
- Some hydrogen removed during the citric acid cycle is picked up by FAD and NAD; one molecule of ATP, three mole- cules of NADH, and one molecule of FADH2 are produced during one complete cycle. Energy associated with electrons and/or with hydrogen picked up by NAD and FAD is gradually released as the electrons are passed along the electron transport system; some of this energy is transferred to ATP molecules during oxidative phosphorylation.
- Energy used in ATP synthesis during oxidative phosphor-ylation is believed to be derived from a gradient of protons formed across the inner membrane of a mitochondrion, while electrons are moving in the electron transport system by chemiosmosis.
- Altogether, 38 ATP molecules are produced during the complete aerobic respiration of one glucose molecule; two are used to prime the process, so there is a net gain of 36 ATP molecules.
- In addition to photosynthesis and respiration, other metabolic pathways are required for growth, development, reproduc- tion, and survival. Essential products of additional pathways include nucleotides, proteins, chlorophylls, and fatty acids. Secondary metabolites include alkaloids, phenolics, and terpenoids.
- Conversion of sugar produced by photosynthesis to fats, proteins, complex carbohydrates, and other substances is termed assimilation. Digestion takes place within plant cells with the aid of enzymes. During digestion, large insoluble mole- cules are broken down by hydrolysis to smaller soluble forms that can be transported to other parts of the plant.