
Plant Biology-Cells

CELL STRUCTURE AND COMMUNICATION
Plant cells typically have a cell wall surrounding the protoplasm, which consists of all the living components of a cell. These living components are bounded by a membrane called the plasma membrane. All cellular components between the plasma membrane and a relatively large body called the nucleus are known as cytoplasm. Within the cytoplasm is a souplike fluid called cytosol, in which various bodies called organelles are dispersed. Organelles are persistent structures of various shapes and sizes with specialized functions in the cell; most, but not all, are bounded by membranes
playlist- Biochemistry – Carbohydrates, Lipids and Cell Membrane
video- Introduction to Carbohydrates (stored plant energy)
video- Polysaccharides (Glycogen, Starch and Cellulose)
The Cell Wall
The main structural component of cell walls is cellulose, which is composed of 100 to 15,000 glucose monomers in long chains, and is the most abundant polymer on earth. As a primary food source for grazing animals and at least indirectly for nearly all other living organisms, it could be said that most life on earth relies directly or indirectly on the cell wall. Humans also depend on cell walls because they provide clothing, shelter, furniture, paper, and fuel.
In addition to cellulose, cell walls typically contain a matrix of hemicellulose (a gluelike substance that holds cellulose fibrils together), pectin (the organic material that gives stiffness to fruit jellies), and glycoproteins (proteins that have sugars associated with their molecules). A middle lamella, which consists of a layer of pectin, is first produced when new cell walls are formed. This middle lamella is normally shared by two adjacent cells and is so thin that it may not be visible with an ordinary light microscope unless it is specially stained. A flexible primary wall, consisting of a fine network of cellulose, hemicellulose, pectin, and glycoproteins, is laid down on either side of the middle lamella Reorganization, synthesis of new molecules, and insertion of new wall polymers lead to – rearrangement of the cell wall during growth. Secondary walls, which are produced inside the primary walls, are derived from primary walls by thickening and inclusion of lignin, a complex polymer. Secondary cell walls of plants generally contain more cellulose (40% to 80%) than primary walls. As the cell ages, wall thickness can vary, occupying as little as 5% to more than 95% of the volume of the cells. During secondary wall formation, cellulose microfibrils become embedded in lignin, much like steel rods are embedded in concrete to form prestressed concrete
The Plasma Membrane
The outer boundary of the living part of the cell, the plasma membrane, [is] composed of phospholipids arranged in two layers, with proteins interspersed throughout This fluid mosaic model for the plasma membrane implies a dynamic structure with numerous components, some of which can migrate and interact directly with each other. Covalent bonds link carbohydrates to both lipids and proteins on the outer surfaces of membranes
videos- Fatty Acids, Glycerol, and Lipids, Proteins-Aminos
video– Cell Membrane and Fluid Mosaic Model
The Nucleus
The nucleus is the control center of the cell Each nucleus is bounded by two membranes, which together constitute the nuclear envelope. The nucleus contains a granular-appearing fluid called nucleoplasm, which is packed with short fibers that are about 10 nanometers in diameter; several different larger bodies are suspended within it. Of the larger nuclear bodies, the most noticeable are one or more nucleoli (singular: nucleolus), which are composed primarily of RNA and associated proteins. The nucleus includes thin strands of chromatin. When a nucleus divides, the chromatin strands coil, becoming shorter and thicker, and in their condensed condition, they are called chromosomes. Chromatin is composed of protein and DNA
The Endoplasmic Reticulum
The outer membrane of the nucleus is connected and continuous with the endoplasmic reticulum. The endoplasmic reticulum facilitates cellular communication and channeling of materials.
Videos– Endoplasmic Reticulum and Golgi Apparatus, Plant Cell Organelles, Translation: Initiation and Ribosomes
Ribosomes
Ribosomal subunits are assembled within the nucleolus, released, and in association with special RNA molecules, they initiate protein synthesis. Once assembled, complete ribosomes may line the outside of the endoplasmic reticulum but can also occur unattached in the cytoplasm, chloroplasts, or other organelles.
Dictyosomes (Cisterna)
Stacks of flattened discs or vesicles known as dictyosomes may be scattered throughout the cytoplasm of a cell. Dictyosomes are often bounded by branching tubules that originate from the endoplasmic reticulum, but are not directly connected to it Dictyosomes are involved in the modification of carbohydrates attached to proteins that are synthesized and packaged in the endoplasmic reticulum. Complex polysaccharides are also assembled within the dictyosomes and collect in small vesicles (tiny, blisterlike bodies) that are pinched off from the margins. These vesicles migrate to the plasma membrane, fuse with it, and secrete their contents outside of the cell. Substances secreted by vesicles may include cell-wall polysaccharides, floral nectars, and essential oils found in herbs.
Plastids
Most living plant cells have several kinds of plastids, with the chloroplasts
Seventy-five to 125 is quite common, with the green cells of a few plants having up to several hundred.
Within the chloroplast are numerous grana (singular: granum), which are formed from membranes and have the appearance of stacks of coins with double membranes. There are usually about 40 to 60 grana linked together by arms in each chloroplast, and each granum may contain from two or three to more than 100 stacked thylakoids. In reality, thylakoids are part of an overlapping and continuous membrane system suspended in the liquid portion of the chloroplast
The thylakoid membranes contain green chlorophyll and other pigments. These “coin stacks” of grana are vital to life as we know it, for it is within the thylakoids that the first steps of the important process of photosynthesis The liquid portion of the chloroplast is a colorless fluid matrix called stroma, which contains enzymes involved in photosynthesis.
Chromoplasts are another type of plastid found in some cells of more complex plants. Although chromoplasts are similar to chloroplasts in size, they vary considerably in shape, often being somewhat angular. They sometimes develop from chloroplasts through internal changes that include the disappearance of chlorophyll. Chromoplasts are yellow, orange, or red in color due to the presence of carotenoid pigments, which they synthesize and accumulate. They are most abundant in the yellow, orange, or some red parts of plants, such as ripe tomatoes, carrots, or red peppers These carotenoid pigments, which are lipid soluble, are not, however, the predominant pigments in most red flower petals. The anthocyanin pigments of most red flower petals are water soluble.
Leucoplasts are yet another type of plastid common to cells of higher plants. They are essentially colorless and include amyloplasts, which synthesize starches, and elaioplasts, which synthesize oils. If exposed to light, some leucoplasts will develop into chloroplasts, and vice versa.
Plastids of all types develop from proplastids
.
Mitochondria
Mitochondria (singular: mitochondrion) are often referred to as the powerhouses of the cell, for it is within them that energy is released from organic molecules by the process of cellular respiration
Carbon skeletons and fatty acid chains are also rearranged within mitochondria, allowing for the building of a wide variety of organic molecules.
Microbodies
Various small bodies distributed throughout the cytoplasm tend to give it a granular appearance. Examples of such components include types of small, spherical organelles called microbodies, which contain specialized enzymes and are bounded by a single membrane. Peroxisomes, for instance, contain enzymes needed by some plants to survive during hot conditions in a process called photorespiration
Vacuoles
In a mature living plant cell, as much as 90% or more of the volume may be taken up by one or two large central vacuoles that are bounded by vacuolar membranes (tonoplasts) The vacuolar membranes, which constitute the inner boundaries of the living part of the cell, are similar in structure and function
to plasma membranes.
The Cytoskeleton
The cytoskeleton is involved in movement within a cell and in a cell’s architecture. It is an intricate network con-structed mainly of two kinds of fibers—microtubules and –microfilaments.
Microtubules control the addition of cell-ulose to the cell wall . They are also involved in cell division, movement of cytoplasmic organelles, controlling the movement of vesicles containing cell-wall components assembled by dictyosomes, and movement of the tiny whiplike flagella and cilia possessed by some cells Microtubules are unbranched, thin, hollow, tubelike structures that resemble tiny straws. They are composed of proteins called tubulins and are of varying lengths, most being between 15 and 25 nanometers in diameter. They are most commonly found just inside the plasma membrane. Microtubules are also found in the special fibers that form the spindles and phragmoplasts of dividing cells
Microfilaments, which play a major role in the contraction and movement of cells in multicellular animals, are present in nearly all cells. They are three or four times thinner than microtubules and consist of long, fine threads of protein with an average diameter of 6 nanometers. They are often in bundles and appear to play a role in the cytoplasmic streaming (sometimes referred to as cyclosis) that occurs in all living cells. When cytoplasmic streaming is occurring, a microscope reveals the apparent movement of organelles as a current within the cytoplasm carries them around within the walls. This streaming probably facilitates exchanges of materials within the cell and plays a role in the movement of substances from cell to cell. The precise nature and origin of cytoplasmic streaming is still not known, but there is evidence that bundles of microfilaments may be responsible for it. Other evidence suggests that it may be related to the transport of cellular substances by microtubules.
CELLULAR REPRODUCTION
The Cell Cycle
When cells divide, they go through an orderly series of events known as the cell cycle. This cycle is usually divided into interphase and mitosis, mitosis itself being subdivided into four phases
videos- Cell Cycle and Interphase, Cytokinesis: Plant vs. Animal Cells, phases of mitosis
Interphase
Living cells that are not dividing are said to be in interphase, a period during which chromosomes are not visible with light microscopes. It is such cells that have been discussed up to this point. For many years, immature cells were considered to be “resting” when they were not actually dividing, but we know now that three consecutive periods of intense activity take place during interphase. These intervals are designated as gap (or growth) 1, synthesis, and gap (or growth) 2 periods, usually referred to as G1, S, and G2, respectively.
Also, ribosomes, RNA, and substances that either inhibit or stimulate the S period that follows are produced. During the S period, the unique process of DNA replication (duplication) takes place. In the G2 period, mitochondria and other organelles divide, and microtubules and other substances directly involved in mitosis are produced. Coiling and condensation of chromosomes also begin during G2.
Mitosis
Strictly speaking, mitosis refers to the division of the nucleus alone, but with a few exceptions seen in algae and fungi the division of the remainder of the cell, called cytokinesis, normally accompanies or follows mitosis. Both processes will be considered together here.
In flowering plants, conifers, and other higher plants, mitosis occurs in specific regions, or tissues, called meri-stems ( Meristems are found in the root and stem tips and also in a thin, perforated, and branching cylinder of tissue called the vascular cambium (often referred to simply as the cambium), located in the interior of some stems and roots a short distance from the surface. In some herbaceous and most woody plants, a second meristem similar in form to the cambium lies between the cambium and the outer bark. This second meristem is called the cork cambium
Mitosis is initiated with the appearance of a ringlike preprophase band of microtubules just beneath the plasma membrane and isusually divided into four arbitrary phases
Prophase
The main features of prophase are (1) the chromosomes become shorter and thicker, and their two-stranded nature becomes apparent; (2) the nuclear envelope fragments, and the nucleolus disintegrates.
. Metaphase
The main feature of metaphase is the alignment of the chromosomes in a circle midway between the two poles around the circumference of the spindle and in the same plane as that previously occupied by the preprophase band. This invisible circular plate is perpendicular to the axis of the spindle and is something like the equator of a globe.
Anaphase
Anaphase—the briefest of the phases—involves the sister chromatids of each chromosome separating and moving to opposite poles
Telophase
The five main features of telophase are (1) each group of daughter chromosomes becomes surrounded by a reformed nuclear envelope; (2) the daughter chromosomes become longer and thinner and finally become indistinguishable; (3) nucleoli reappear; (4) many of the spindle fibers disintegrate; and (5) a cell plate forms.
HIGHER PLANT CELLS VERSUS ANIMAL CELLS
All animals have either internal or external skeletons or skeleton-like systems to support their tissues. Animal cells do not have cell walls; instead, the plasma membrane, called the cell membrane by most zoologists (animal scientists), forms the outer boundary of animal cells. Higher plant cells have walls that are thickened and rigid to varying degrees, with a framework of cellulose fibrils. Higher plant cells also have plasmodesmata connecting the protoplasts with each other through microscopic holes in the walls. Animal cells lack plasmodesmata since they have no walls. When higher plant cells divide, a cell plate is formed during the telophase of mitosis, but cell plates do not form in animal cells, which divide by pinching in two.
Summary
- All living organisms are composed of cells. Cells are modified according to the functions they perform; some live for a few days, while others live for many years.
- The discovery of cells is associated with the development of the microscope. In 1665, Robert Hooke coined the word cells for boxlike compartments he saw in cork. Leeuwenhoek and Grew reported frequently during the next 50 years on the existence ofcells in a variety of tissues.
- In 1809, Lamarck concluded that all living tissue is composed of cells, and in 1824, Dutrochet reinforced Lamarck’s conclusions. In 1833, Brown discovered that all cells contain a nucleus, and shortly thereafter, Schleiden saw a nucleolus within a nucleus. Schleiden and Schwann are credited with developing the cell theory in 1838 to 1839. The theory holds that all living organisms are composed of cells and that cells form a unifying, structural basis of organization.
- In 1858, Virchow contended that every cell comes from a preexisting cell and that there is no spontaneous generation of cells from dust. In 1862, Pasteur experimentally confirmed Virchow’s contentions and later proved that fermentation involves activity of yeast cells. In 1897, Buchner found that yeast cells do not need to be alive for fermentation to occur. This led to the discovery of enzymes.
- Light microscopes can magnify up to 1,500 times. Thinly sliced materials can be viewed with compound microscopes. Opaque objects can be viewed with stereo-microscopes; most magnify up to 30 times.
- Electron microscopes have electromagnetic lenses and a beam of electrons within a vacuum that achieve magnification. Transmission electron microscopes magnify up to 200,000 or more times. Scanning electron microscopes, which can be used with opaque objects, usually magnify up to 10,000 times. 7. Scanning tunneling microscopes use a minute probe to scan surfaces at a width as narrow as that of two atoms.
- Eukaryotic cells are the subject of this topc. Pro-karyotic cells, lack some of the features of eukaryotic cell.
- Cells are minute, varying in diameter between 10 and 100 micrometers. They number into the billions in larger organisms, such as trees. Plant cells are bounded by walls that surround the living protoplast. The cytoplasm contains a souplike fluid called the cytoso and all cellular components between the plasma membrane and nucleus.
- A pectic middle lamella is sandwiched between the primary cell walls of adjacent cells. The primary wall and also the secondary cell wall, often added inside the primary wall, are composed of cellulose polymers, with hemicelluloses and glycoproteins. Secondary cell walls contain lignin, which strengthens the wall.
- Living cells are in contact with one another via fine strands of cytoplasm called plasmodesmata, which often extend through minute holes in the walls.
- A flexible plasma membrane, which is sandwich-like and often forms folds, constitutes the outer boundary of the cytoplasm. It regulates the substances that enter and leave the cell.
- The nucleus is bounded by a nuclear envelope consisting of two membranes that are perforated by numerous pores. Within the nucleus are a fluid called nucleoplasm, one or more spherical nucleoli, and thin strands of chromatin, which condense and become chromosomes when nuclei divide. Each species of organism has a specific number of chromosomes in each cell.
- The endoplasmic reticulum is a system of flattened sacs and tubes associated with the storing and transporting of protein and other cell products. Granular particles called ribosomes, which function in protein synthesis, may line the outer surfaces of the endoplasmic reticulum. Ribosomes also occur independently in the cytoplasm.
- Dictyosomes are structures that appear as stacks of sacs and function as collecting and packaging centers for the cell.
- Plastids are larger green, orange, red, or colorless organelles. Green plastids, known as chloroplasts, contain enzymes that catalyze reactions of photosynthesis. These reactions take place in the membranes of structures that resemble stacks of coins, called thylakoids, as well as the surrounding matrix, called the stroma. Plastids develop from proplastids, which divide frequently, and also arise from the division of mature plastids.
- Mitochondria are tiny, numerous organelles that are bounded by two membranes with inner platelike folds called cristae; they are associatedwith cellular respiration.
- One or more vacuoles may occupy 90% or more of the volume of a mature cell. Vacuoles are bounded by a vacuolar membrane (tonoplast) and contain a watery fluid called cell sap. Cell sap contains dissolved substances and sometimes water-soluble red or blue anthocyanin pigments.
- The cytoskeleton, which is involved in the architecture of cells and internal movement, is composed of microtubules and microfilaments. Microfilaments may be responsible for cytoplasmic streaming.
- Cells that are not dividing are in interphase, which is subdivided into three periods of intense activity that precede mitosis or division of the nucleus. Mitosis is usually accompanied by division of the rest of the cell and takes place in meristems.
- Mitosis is arbitrarily divided into four phases: (1) prophase, in which the chromosomes and their two-stranded nature become apparent and the nuclear envelope breaks down; (2) metaphase, in which the chromosomes become aligned at the equator of the cell; a spindle composed of spindle fibers is fully developed, with some spindle fibers being attached to the chromosomes at their centromeres; (3) anaphase, in which the sister chromatids of each chromosome (now called daughter chromosomes) separate lengthwise, with each group of daughter chromosomes migrating to opposite poles of the cell; and (4) telophase, in which each group of daughter chromosomes becomes surrounded by a nuclear envelope, thus becoming new nuclei, and a wall dividing the daughter nuclei forms, creating two daughter cells.
- Animal cells differ from those of higher plants in not having a wall, plastids, or large vacuoles. Also, they have keg-shaped centrioles in pairs just outside the nucleus and pinch in two instead of forming a cell plate when they divide.
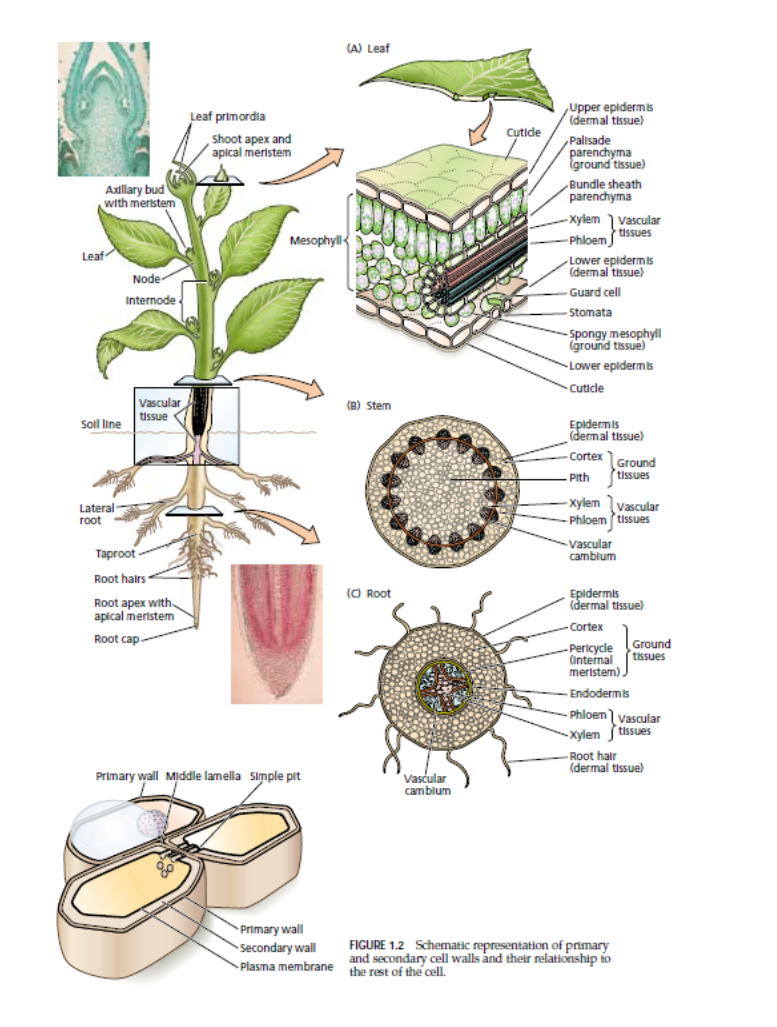
Tissues
MERISTEMATIC TISSUES
Unlike animals, plants have permanent regions of growth called meristems, or meristematic tissues, where cells actively divide
Apical Meristems
Apical meristems are meristematic tissues found at, or near, the tips of roots and shoots, which increase in length as the apical meristems produce new cells. This type of growth is known as primary growth. Three primary meristems, as well as embryo leaves and buds, develop from apical meristems. These primary meristems are called protoderm, ground meristem, and procambium. The tissues they produce are called primary tissues.
Lateral Meristems
The vascular cambium and cork cambium, discussed next, are lateral meristems, which produce tissues that increase the girth of roots and stems. Such growth is termed secondary growth.
Vascular Cambium
The vascular cambium, often referred to simply as the cambium, produces secondary tissues that function primarily in support and conduction. The cambium, which extends throughout the length of roots and stems in perennial and many annual plants, is in the form of a thin cylinder of mostly brick-shaped cells. The cambial cylinder often branches, except at the tips, and the tissues it produces are responsible for most of the increase in a plant’s girth as it grows. The individual remaining cells of the cambium are referred to as initials, while their sister cells are called derivatives.
Cork Cambium
The cork cambium, like the vascular cambium, is in the form of a thin cylinder that runs the length of roots and stems of woody plants. It lies outside of the vascular cambium, just inside the outer bark, which it produces. The tissues laid down by the vascular cambium and the cork cambium are called secondary tissues, since they are produced after the primary tissues have matured
Intercalary Meristems
Grasses and related plants have neither a vascular cambium nor a cork cambium. They do, however, have apical meri-stems, and, in the vicinity of nodes (leaf attachment areas), they have other meristematic tissues called intercalary meristems. The intercalary meristems develop at intervals along stems, where, like the tissues produced by apical meristems, their tissues add to stem length.
Simple Tissues
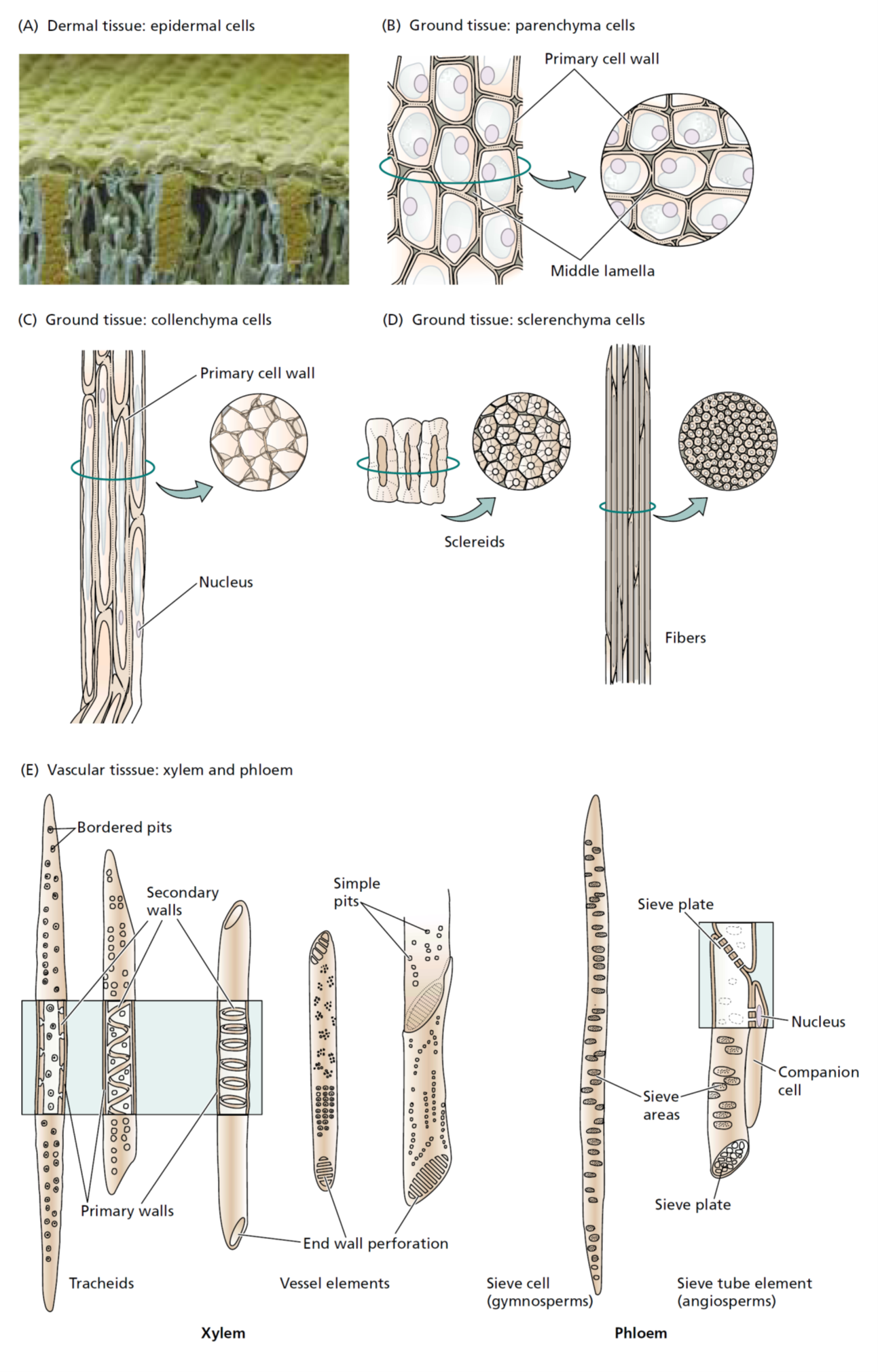
Parenchyma
Parenchyma tissue is composed of parenchyma cells ( They are more or less spherical in shape when they are first produced, but when all the parenchyma cells push up against one another, their thin, pliable walls are flattened at the points of contact. As a result, parenchyma cells assume various shapes and sizes, with the majority having 14 sides. They tend to have large vacuoles and may contain starch grains, oils, tannins (tanning or dyeing substances), crystals, and various other secretions. More often than not, parenchyma cells have spaces between them; in fact, in water lilies and other aquatic plants, the intercellular spaces are quite extensive and form a network throughout the entire plant. This type of parenchyma tissue—with extensive connected air spaces—is referred to as aerenchyma.
Parenchyma cells containing numerous chloroplasts (as found in leaves) are collectively referred to as chlorenchyma tissue. Chlorenchyma tissues function mainly in photosynthesis, while parenchyma tissues without chloroplasts function mostly in food or water storage. For example, the soft, edible parts of most fruits and vegetables consist largely of parenchyma.
Collenchyma
Collenchyma cells like parenchyma cells, have living cytoplasm and may remain alive a long time. Their walls generally are thicker and more uneven in thickness than those of parenchyma cells. The unevenness is due to extra primary wall in the corners. Collenchyma cells often occur just beneath the epidermis; typically, they are longer than they are wide, and their walls are pliable as well as strong. They provide flexible support for both growing organs and mature organs, such
as leaves and floral parts. The “strings” of celery that get stuck in our teeth, for example, are composed of collenchyma cells
Sclerenchyma
Sclerenchyma tissue consists of cells that have thick, tough, secondary walls, normally impregnated with lignin. Most sclerenchyma cells are dead at maturity and function in support. Two forms of sclerenchyma occur: sclereids and fibers. Sclereids may be randomly distributed in other tissues. For example, the slightly gritty texture of pears is due to the presence of groups of sclereids, or stone cells, as they are sometimes called.
Xylem
Xylem tissue is an important component of the “plumbing” and storage systems of a plant and is the chief conducting tissue throughout all organs for water and minerals absorbed by the roots. Xylem consists of a combination of parenchyma cells, fibers, vessels, tracheids, and ray cells (Vessels are long tubes composed of individual cells called vessel elements that are open at each end. As each vessel element develops, the perforation plate, in some instances, can become barlike strips of wall material that extend across the openings. However, the flow of fluid through the vessels is not blocked by the strips.
Tracheids, which, like vessel elements, are dead at maturity and have relatively thick secondary cell walls, are tapered at each end, the ends overlapping with those of other tracheids. Tracheids have no openings similar to those of vessels, but there are usually pairs of pits present wherever two tracheids are in contact with one another. Pits are areas in which no secondary wall material has been deposited and, they allow water to pass from cell to cell. Figure 4.8 illustrates how, in some plants, pit pairs function in regulating the passage of water between adjacent cells. In cone-bearing trees and certain other non-flowering plants, the xylem is composed almost entirely of tracheids. The walls of many tracheids, as well as vessel elements, have spiral thickenings on them that are easily seen with the light microscope Most conduction through xylem is upward, but some is lateral (sideways). The lateral conduction takes place in the rays. Ray cells, which also function in food storage, are actually long-lived parenchyma cells that are produced in horizontal rows by special ray initials of the vascular cambium. In woody plants, the rays radiate out from the center of stems and roots like the spokes of a wheel.
Phloem
Phloem tissue which conducts dissolved food materials (primarily sugars) produced by photosynthesis throughout the plant, is composed mostly of two types of cells without secondary walls. The relatively large, more or less cylindrical sieve tube members have narrower, more tapered companion cells closely associated with them. Phloem is derived from the parent cells of the cambium, which also produce xylem cells; it often also includes fibers, parenchyma, and ray cells. Sieve tube members, like vessel elements, are laid end to end, forming sieve tubes. Unlike vessel elements, however, the end walls have no large openings; instead, the walls are full of small pores through which the cytoplasm extends from cell to cell. These porous regions of sieve tube members are called sieve plates. Sieve tube members have no nuclei at maturity, even though their cytoplasm is very active in the conduction of food materials in solution throughout the plant. Apparently, the adjacent companion cells form a very close relationship with the sieve tubes next to them and aid in the conduction of the food. Living sieve tube members contain a polymer called callose that stays in solution as long as the cell contents are under pressure. If an insect such as an aphid injures a cell, however, the pressure drops, and the callose precipitates. The callose and a phloem protein are then carried to the nearest sieve plate where they form a callus plug that prevents leaking of the sieve tube contents.
Sieve cells, which are found in ferns and cone-bearing trees, are similar to sieve tube members but tend to overlap at their ends rather than form continuous tubes. Like sieve tube members, they have no nuclei at maturity, but they have no adjacent companion cells. They do have adjacent albuminous cells, which are equivalent to companion cells and apparently function in the same manner.
Videos- Plant Nutrition and Transport, Water transport in plants, Transport in Phloem, Plant Structure.
Epidermis
The outermost layer of cells of all young plant organs is called the epidermis. Since it is in direct contact with the environment, it is subject to modification by the environment and often includes several different kinds of cells. The epidermis is usually one cell thick, but a few plants produce aerial roots called velamen roots (e.g., orchids) in which the epidermis may be several cells thick, with the outer cells functioning something like a sponge. Such a multiple-layered epidermis also occurs in the leaves of some tropical figs and members of the Pepper Family (Piperaceae), where it protects a plant from desiccation. Most epidermal cells secrete a fatty substance called cutin within and on the surface of the outer walls. Cutin forms a protective layer called the cuticle The thickness of the cuticle (or, more importantly, wax secreted on top of the cuticle by the epidermis) to a large extent determines how much water is lost through the cell walls by evaporation. The cuticle is also exceptionally resistant to bacteria and other disease organisms and has been recovered from fossil plants millions of years old. The waxes deposited on the cuticle in a number of plants can reach the surface by diffusion, migrate between cells, or travel through microscopic channels in the cell walls. The susceptibility of a plant to herbicides may depend on the thickness of these wax layers. Some wax deposits are extensive enough to have commercial value. Carnauba wax, for example, is deposited on the leaves of the wax palm. It and other waxes are harvested for use in polishes and, in the past, for phonograph records. In colonial times, a wax obtained from boiling leaves and fruits of the wax myrtle was used to make bayberry candles. In leaves, the epidermal cell walls perpendicular to the surface often assume bizarre shapes that, under the microscope, give them the appearance of pieces of a jigsaw puzzle. Epidermal cells of roots produce tubular extensions called root hairs a short distance behind the growing tips. The root hairs greatly increase the absorptive area of the surface. Hairs of a different nature occur on the epidermis of above-ground parts of plants. These hairs form outgrowths consisting of one to several cells Leaves also have numerous small pores, the stomata, bordered by pairs of specialized epidermal cells called guard cells Guard cells differ in shape from other epidermal cells; they also differ in that chloroplasts are present within themSome epidermal cells may be modified as glands that secrete protective or other substances, or modified as hairs that either reduce water loss or repel insects and animals that might otherwise consume them .
Periderm
In woody plants, the epidermis is sloughed off and replaced by a periderm after the cork cambium begins producing new tissues that increase the girth of the stem or root. The peri-derm constitutes the outer bark and is primarily composed of somewhat rectangular and boxlike cork cells, which are dead at maturity While the cytoplasm of cork cells is still functioning, it secretes a fatty substance, suberin, into the walls. This makes cork cells waterproof and helps them protect the phloem and other tissues beneath the bark from drying out, mechanical injury, and freezing temperatures. Some cork tissues, such as those produced by the cork oak, are harvested commercially and are used for bottle corks and in the manufacture of linoleum and gaskets. Some parts of a cork cambium form pockets of loosely arranged parenchyma cells that are not impregnated with suberin. These pockets of tissue protrude through the surface of the periderm; they are called and function in gas exchange between the air and the interior of the stem. The fissures in the bark of trees have lenticels at their bases. The various tissues discussed are shown as they occur in a woody stem in Figure 6.6.
Secretory Cells and Tissues
All cells secrete certain substances that can damage the cytoplasm, if allowed to accumulate internally. Such materials either must be isolated from the cytoplasm of the cells in which they originate or moved outside of the plant body. Often, the substances consist of waste products that are of no further use to the plant, but some substances, such as nectar, perfumes, and plant hormones, are vital to normal plant functions. Secretory cells may function individually or as part of a secretory tissue. Secretory cells or tissues, which often are derived from parenchyma, can occur in a wide variety of places in a plant. Among the most common secretory tissues are those that secrete nectar in flowers; oils in citrus, mint, and many other leaves; mucilage in the glandular hairs of sundews and other insect-trapping plants; latex in members of several plant families, such as the Spurge Family; and resins in coniferous plants, such as pine trees. Latex and resins are usually secreted by cells lining tubelike ducts that form networks throughout certain plant species Some plant secretions, such as pine resin, rubber, mint oil, and opium, have considerable commercial value.
Summary
- A group of cells performing a common function is called a tissue. Apical meristems are found in the vicinity of the tips of roots and stems; the vascular cambium and the cork cambium occur as lengthwise cylinders within roots and stems; intercalary meristems occur in the vicinity of nodes of grasses and related plants.
- Tissues produced by meristems consist of one to several kinds of cells. They include parenchyma, collenchyma, sclerenchyma, epidermis, xylem, phloem, periderm, and secretory tissues.
- Parenchyma cells are thin-walled, while collenchyma cells have unevenly thickened walls that provide flexible support for various plant organs.
- Two types of sclerenchyma occur—fibers (which are long and tapering) and sclereids (which are short in length); both types have thick walls and are usually dead at maturity.
- Complex tissues have more than one kind of cell. The principal types are xylem, phloem, epidermis, and -periderm.
- Xylem conducts water and minerals throughout the plant. It consists of a combination of parenchyma, fibers, vessels (tubular channels), tracheids (cells with tapering end walls that overlap), and ray cells (involved in lateral conduction).
- Phloem conducts primarily dissolved sugars throughout the plant. It is composed of sieve tubes (made up of cells called sieve tube members), companion cells (that apparently regulate adjacent sieve tube members), arenchyma, ray cells, and fibers. Callose aids in plugging injured sieve tubes. Sieve cells, which have overlapping end walls, and adjacent albuminous
- cells take the place of sieve tube members and companion cells in ferns and cone-bearing trees.
- he epidermis is usually one cell thick, with fatty cutin (forming the cuticle) within and on the surface of the outer walls.
- The epidermis may include guard cells that border pores called stomata; root hairs, which are tubular extensions of single cells; other hairs that consist of one to several cells; and glands that secrete protective substances.
- Periderm, which consists of cork cells and loosely arranged groups of cells comprising lenticels involved in gas exchange,constitutes the outer bark of woody plants.
- Secretory tissues occur in various places in plants; they secrete substances such as nectar, oils, mucilage, latex, and resins.